

Oddělení: Sinice – Cyanobacteria (syn. Cyanophyta ...
Oddělení: Sinice – Cyanobacteria (syn. Cyanophyta ...
Oddělení: Sinice – Cyanobacteria (syn. Cyanophyta ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
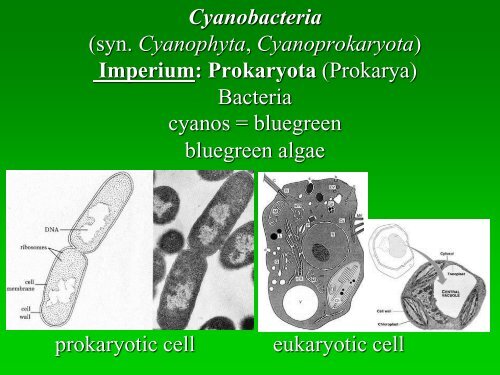
<strong>Cyanobacteria</strong><br />
(<strong>syn</strong>. <strong>Cyanophyta</strong>, Cyanoprokaryota)<br />
Imperium: Prokaryota (Prokarya)<br />
Bacteria<br />
cyanos = bluegreen<br />
bluegreen algae<br />
prokaryotic cell eukaryotic cell
Principal characteristics<br />
coccoid (free living or colonial) or filamentous<br />
thylakoids posses photosystem I and II (also respiration<br />
chain on thylakoids)<br />
thylakoids separated or in lamelas (Gleobacter without<br />
thylak. <strong>–</strong> PS on plasmatic membrane)<br />
chlorophyll a (but also b, c or d)<br />
chromatic adaptation, phycobilisomes<br />
reserve substances <strong>–</strong> cyanophycean starch; cyanophycin<br />
granules (N) and polyphosphate granules (P)<br />
some are able to fix atmospheric nitrogen<br />
ubiquitous<br />
static evolution<br />
asexual reproduction by cell division<br />
horizontal gene transfer (plasmids)
<strong>Cyanobacteria</strong> = G - bakteria<br />
Cell wall structure<br />
Lipopolysacharidy<br />
murein
the cell wall composed of peptidoglycan (murein) similarily to<br />
the other G - bacteria<br />
CW cleaved by lysozym, penicillin prevent CW <strong>syn</strong>thesis<br />
2 sugar derivatives
<strong>Cyanobacteria</strong> = G - bacteria<br />
Arg; Asp<br />
N-reserve
Aerotops (gas vesicles)<br />
Gas vesicles are spindle-shaped structures provides buoancy to these<br />
cells by decreasing their overall cell density, glycoproteins<br />
(impermeable to water but permeable to most gases)<br />
• high light? increased PS → accumulation of polysaccharides → turgor<br />
increases → collaps of vesicles → the cell sinks out<br />
• low light? decreased PS → metabolism of polysaccharides → turgor<br />
decreases → vesicle production → the cell rises up<br />
Microcystis<br />
Tolypothrix, Calothrix <strong>–</strong> gas vesicules only in hormogonia <strong>–</strong> dispersal??
Reserve substances<br />
cyanophycean starch <strong>–</strong> small granules between thylakoids,<br />
-1,4 linked glucan<br />
cyanophycin granules (obr. - CY) <strong>–</strong> polymers of Arg, Asp<br />
polyphosphate granules (volutin) <strong>–</strong> highly polymerized<br />
polyphosphate<br />
Oscillatoria - polyphosphate granules in<br />
the hormogonium
Caboxysomes<br />
(polyhedral bodies) <strong>–</strong> Rubisco: CO 2 polysaccharides<br />
Enclosed in a proteinaceus cover, reserves of the enzymes<br />
RubisCO and carbonic anhydrase (HCO 3 - CO2)<br />
Proteinaceus cover of the carboxysome increases<br />
carboxylase activity of the RubisCO.<br />
Hypothesis: the enzym enclosed in the carboxysome is forced<br />
to the conformation, that prefers CO 2 to O 2.
Pigments<br />
phycobiliproteins<br />
phycobiliproteins fill the „gap“ in the absorbance<br />
spectrum of chlorophylls and carotenoids<br />
major pigments - Cha and phycobiliproteins
Light absorption by phycobiliproteins<br />
Blue, red: Chlorophyll a, (b,d)<br />
- caroten (much higher proportion than<br />
in euk. algae), echinenon, myxoxanthophyll<br />
green (500-600 nm): C-phycoerythrin *<br />
green - yellow (550-650 nm): C-phycocyanin<br />
orange - red (600-675 nm): allophycocyanin*<br />
phycoerythrocyanin (568 nm)<br />
combining of the pigments <strong>–</strong> absorption<br />
in the whole spectrum of visible light<br />
Absorption of light in<br />
a water column
Phycobilisome <strong>–</strong> light harvesting<br />
antenna<br />
for photosystem II and nitrogen reserve
Chromatic adaptation : the relative quantities of phycoerythrin and<br />
phycocyanin determines the color of the cell, dependent<br />
green light phycoerythrin phycocyanin<br />
red light phycocyanin phycoerythrin
Different photo<strong>syn</strong>t. apparatus<br />
Gloeobacter violaceus <strong>–</strong> primitive cyanobacteria, no thylakoids<br />
phycobilisomes placed on plasmatic membrane, isolated from<br />
limestone, Switzerland<br />
former PROCHLOROPHYTES:<br />
no phycobiliproteins (neither phycobilisomes) posses additional<br />
pigments: chl. b, c, d; - caroten, zeaxanthin<br />
thylakoids organized in lamelae, lack of cyanophycin granules
Prochlorophyta <strong>–</strong> polyphyletic group<br />
Acaryochloris marina<br />
distantly related to other cyanobacteria
Prochlorophyta <strong>–</strong> polyphyletic group<br />
Prochloron didemni - Prochlorons are distributed<br />
within the tissues of didemnid ascidians, mainly<br />
in the cloacal cavities under the upper tunic,<br />
Pacific ocean, chlorophyll b<br />
Prochlorothrix hollandica <strong>–</strong> filamentous, in a<br />
brakish Lake Loosdrecht in Holand, masive mats<br />
Prochlorococcus marinus <strong>–</strong> marine picoplankton,<br />
omportant primary producer, chlorophyll a and b<br />
Acaryochloris marina <strong>–</strong> chlorophyll d<br />
Prochloron Prochlorococcus
400 BP of 16 S RNA sequences NJ<br />
Münchhoff et al.,2006<br />
stromatolites
Prochloron <strong>–</strong> phylogeny of 30 strains<br />
Münchhoff et al.,2006<br />
• all strains closely related, forming a single taxon<br />
• shown that there is no evident correlation between<br />
Prochloron sp. and its host or geographic origin.<br />
• Prochlorons isolated from stromatolites (probably<br />
nonsymbiotic origin) only 4 strains formed coherent<br />
group on a phylogenetic tree.<br />
• Phylogenetic analysis demonstrated that the<br />
prochlorophytes are a polyphyletic group within the<br />
cyanobacterial radiation, supporting the view that the<br />
ability to <strong>syn</strong>thesize chlorophyll b evolved several<br />
times separately with consequent loss of the ability to<br />
<strong>syn</strong>thesize phycobilins
• Furthermore, due to the low genetic diversity among<br />
Prochloron strains, this genus could be a relatively<br />
recent lineage thus contradicting the view that these<br />
prochlorophytes are descendants of the ancient<br />
organisms that gave rise to chloroplasts<br />
• In contrast, studies of the chlorophyll b <strong>syn</strong>thesis genes<br />
from P. didemni, Prochlorothrix hollandica and several<br />
green chloroplasts indicated a common origin for these<br />
genes in prochlorophytes and chloroplasts<br />
• It seems likely that horizontal gene transfer may<br />
account for the similarity among the chlorophyll b<br />
<strong>syn</strong>thesis genes in these organisms
Facultative phototrophic anaerobes<br />
both photoaerobic and photoanaerobic habitats<br />
electron donors?<br />
aerobic conditions e - donor water (O 2 production)<br />
2 H 2O+ CO 2 [CH 2O] + O 2 + H 2O<br />
anaerobic conditions e - donor hydrogen sulfide (no O 2 production)<br />
only PS I involved<br />
progenitors - purple bacteria -<br />
bacteriochlorophyll<br />
2 H 2S+ CO 2 [CH 2O] + 2S + H 2O
fill an important ecological niche, fluctuation between aerobic<br />
and anaerobic conditions<br />
Oscillatoria limnetica<br />
Solar Lake <strong>–</strong> small hyper<br />
saline lake (eastern coast<br />
of the Sinai peninsula<br />
(Israel)<br />
thermal stratification;<br />
winter <strong>–</strong> high<br />
concentrations of H 2S<br />
near the bottom<br />
Some species obligate autotrophs (=energy obtained by<br />
PS) or heterotrophs (= energy obtained from organic C)
they develope from<br />
vegetative cells<br />
thick-walled, full of<br />
reserve material<br />
may survive up to 86<br />
years in the sediment<br />
Akinetes <strong>–</strong> resting stages<br />
only in heterecyst forming cyanobacteria<br />
Anabaena<br />
Anabaena - klíčení<br />
Aphanizomenon<br />
sink, overwinter akinete differentiation stimulation
N 2 fixation<br />
fixation = N 2 (N≡N) from atmosphere is fixed into<br />
ammonium (NH 4 + )<br />
N 2 + 8 H + + 16 ATP 2 NH 3 + H 2+ 16 ADP + 16 P<br />
renewable energy source<br />
N <strong>–</strong> limiting element in the sea water, amino-acid's <strong>syn</strong>thesis<br />
only cyanobacteria and bacteria are able to fix nitrogen;<br />
cyanobacteria also produce O 2 → nitrogenase is inactivated<br />
by O 2<br />
spatial or temporal separation of both activities (except for<br />
Trichodesmium)<br />
The most metabolically expensive process in biology!!
Spatial separation - heterocysts<br />
Nostocales, Stigonematales<br />
• thick wall, bigger than<br />
vegetative cells<br />
• differentiation from vegetative<br />
cells<br />
• in heterocysts PSII is<br />
lacking nitrogenase<br />
protection from O2<br />
• they do not fix C, C is<br />
transported in a form of<br />
disaccharides from veg. cells<br />
through the pore channel<br />
heterocysts in<br />
distinctive patterns differentiation of heterocysts, apoptosis
Temporal separatoin<br />
Fix N at night, foto<strong>syn</strong>thesis during the day<br />
In the evening phycobiliprotein's degradation PSII switch off O2<br />
production stopped fast nitrogenase <strong>syn</strong>thesis N2 fixation over night<br />
nitrogenase <strong>syn</strong>thesis stopped, the enzyme is disintegrated by oxygen,<br />
that penetrates to cells re<strong>syn</strong>thesis of phycobiliproteins functional<br />
PSII in the morning the photo<strong>syn</strong>thesis may begin again.<br />
Lyngbya aestuarii
Trichodesmium - diazocytes<br />
Trichodesmium <strong>–</strong> sea water filamentous species, N2 fixation<br />
in the light under aerobic conditions <strong>–</strong> specialized cells <strong>–</strong><br />
diazocytes (contain nitrogenase, adjacent to one another)<br />
anti-nifH <strong>–</strong> přítomnost<br />
nitrogenázy ve fluorescenci<br />
Katagnymene
Morfology of cyanobacteria<br />
• row of cells = trichom<br />
• trichom in the slime sheath = filament<br />
• the filament may be composed by more<br />
trichomes<br />
Microcoleus<br />
Phormidium<br />
• mucilage sheath - movement, UV protection, colony<br />
addition, capacity to capture micronutrients
• gliding<br />
Movement<br />
Oscillatoria<br />
• buoancy change <strong>–</strong> gas vesicles<br />
Merismopedia Waden See <strong>–</strong><br />
migrate within the substrate<br />
Cyanothece aeruginosa
Gliding motion<br />
oscillin<br />
junctional pores<br />
junctional pores<br />
orientation<br />
of oscillin<br />
proteins<br />
reversal of<br />
gliding<br />
active movement on a solid substrate (up to 600 m/sec.)
Morphology and cell division<br />
centripetal (Synechoccocus, Cyanothece)<br />
nanocytes (Aphanothece)
Morphology and cell division<br />
endospores-baeocytes<br />
(Cyanocystis)<br />
exocytes, exospores (Chamaesiphon)
division in 2 or 3 directions plate-like colonies (Merismopedia)<br />
or irregular colonies (Microcystis) embedded in a mucilage<br />
M. flos-aque<br />
Morphology and cell division<br />
Merismopedia Microcystis<br />
M. aeruginosa
Morphology and cell division<br />
division of truly filamentous cyanobacteria (trichomes)<br />
<strong>–</strong>hormogonia = short pieces of trichomes, move away<br />
by gliding (trichomes fragmentation or necridia<br />
formation)<br />
Lyngbya<br />
change in environmental parameters, gliding stage<br />
36-48hrs, secretion of mucilage
Trichomes and branching<br />
false branching = trichome breaks within the mucilage sheath<br />
(Scytonema,Tolypothrix)<br />
Scytonema
Trichomes and branching<br />
true branching = change of the plain of the cell division<br />
(Stigonema, Hapalosiphon)
DNA transfer<br />
horizontal gene transfer: ca 10-16% genes obtained<br />
by horizontal transfer by means of plasmids or<br />
viruses (cyanophages)<br />
conjugation not known<br />
cyanophages may serve as a „reservoir“ of genes<br />
for the host <strong>–</strong> enable the host to adapt to the<br />
changing environmental conditions
analyzed 11 genomes of cyanobacteria, 1128 genes; left half <strong>–</strong> congruent<br />
topology (only one the most parsimony tree exists), right half -685 genes -<br />
61% - conflict with majority signal
Example of intraphylum transfer:<br />
a hemolysin-like protein.<br />
(A) Anabaena sp.; (Tr) Trichodesmium<br />
erythraeum; (S) Synechocystis sp.; (1P)<br />
Prochlorococcus marinus; (2P) Prochlorococcus<br />
marinus; (3P) Prochlorococcus marinus; (mS)<br />
marine Synechococcus ; (Th)<br />
Thermo<strong>syn</strong>echococcus elongatus; (G)<br />
Gloeobacter violaceus; (N) Nostoc<br />
punctiforme; (C) Crocosphaera watsonii
Castenholz (1992)<br />
Static evolution<br />
Why the precambric cyanobacteria look like present-day<br />
ones?<br />
• long generation time and larger genome size compared to bacteria<br />
• often polyploidy <strong>–</strong> mutation in a single allele will not be expressed (x<br />
haploid eukaryotes)<br />
• reparation mechanisms are probably involved (shown in a chloroplast)<br />
• low rate of spontaneous mutations (also under mutagenesis)
věk sinic 2,5-0,6<br />
Van den Hoek, s. 36
Stromatolites<br />
stromatolites <strong>–</strong> trapping and binding of the sdiments as well as carbonate<br />
precipitation in cyanobacterial sheaths (from 2.7 mld) or „grain trapping“ <strong>–</strong><br />
Precambrium - 3,5 mld years old<br />
Québec - Kanada
heliotropic <strong>–</strong><br />
grow toward<br />
sunlight<br />
Eoentophysalis<br />
Stromatolites<br />
Paleolyngbia<br />
Oscollatoriopsis<br />
fosil cyanobacteria from the stromatolite, 1.4 mld years old<br />
Counting of the<br />
number of laminae<br />
solar year varied considerably<br />
1billion y ago <strong>–</strong> 435 days
Stromatolites<br />
live stromatolites <strong>–</strong> Western Australia, Shark‘s Bay<br />
hypersaline<br />
2 billion y ago <strong>–</strong> no grazers
Stromatolites<br />
live stromatolites <strong>–</strong> southern Florida, Bahamas<br />
Schizothrix<br />
high tidal current
Heterocystous cyanobacteria phylogeny<br />
recentní fosil akinetes<br />
1,5 mld<br />
1,65 mld<br />
2,1 mld<br />
Tomitani et al. 2006
Heterocystous cyanobacteria phylogeny<br />
Tomitani et al. 2006
The systematics of <strong>Cyanobacteria</strong><br />
Castenholz (1992): only genera, so called static evolution<br />
Van den Hoek et al. (1995): 2000 species in 150 genera<br />
Komárek & Anagnostidis (1999): thousands of species
The systematics of <strong>Cyanobacteria</strong><br />
Taxonomic scheme according to Bergey's Manual of<br />
Systematic Bacteriology 2nd Edition<br />
D.R. BOONE, R.W. CASTENHOLZ, G.M. GARRITY (eds), 2001.<br />
not congruent with a molecular phylogeny<br />
I. Chroococcales <strong>–</strong> single celled, free living or colonial, binary fission<br />
II. Pleurocapsales <strong>–</strong> multiple fission, baeocytes<br />
III. Oscillatoriales <strong>–</strong> simple filamentous cyanobacteria without<br />
heterocysts and akinetes<br />
IV. Nostocales - filamentous cyanobacteria with heterocysts and akinetes,<br />
some of them with false branching<br />
V. Stigonematales - filamentous cyanobacteria with heterocysts and true<br />
branching
Microbial Genome Sequencing<br />
Project, 2007
Tomitani et al.2006
Gloeobacter violaceus<br />
Chroococcales<br />
Synechoccocus <strong>–</strong>picoplanktonic, freshwater/marine, eutrophic,<br />
Synechocystis <strong>–</strong> marine/mineral waters<br />
Synechocystis
Chroococcales<br />
Chroococcus <strong>–</strong> marine/freshwater, soil, damp rocks<br />
Cyanothece <strong>–</strong> peat bogs<br />
C. turgidus<br />
C. limneticus
Chroococcales<br />
Aphanothece <strong>–</strong> mucilage colonies, subaerical<br />
(A. castagnei, lime stone), benthos (A. stagnina, eutrophic ponds)<br />
(A. microscopica, acidic waters)<br />
Gloeocapsa <strong>–</strong> concentrically layered slime, damp rocks (granite)<br />
Aphanothece<br />
A. microscopica<br />
A. castagnei<br />
A. stagnina
Chroococcales<br />
Merismopedia <strong>–</strong> plate-like colonies embeded in mucilage<br />
Chamaesiphon <strong>–</strong> freshwater, pear-shaped cells, stones in fastflowing<br />
streams<br />
Chroococcidiopsis <strong>–</strong> extreme biotopes, NASA<br />
M. angularis<br />
C. rostafinski<br />
C. amathystinus
Chroococcales<br />
Microcystis <strong>–</strong> irregular colonies in mucilage, aerotopes,<br />
only planktonic, cyanotoxines, water blooms
M. ichtyoblabe<br />
M. ichtyoblabe<br />
Chroococcales
Chroococcales
Otsuda et al. 2001 (IJSEM)<br />
proposed to unify these five species into M. aeruginosa under the<br />
Rules of the Bacteriological Code
M. viridis<br />
M. wesenbergii<br />
Chroococcales<br />
M. viridis<br />
M. wesenbergii
Chroococcales<br />
Woronichinia <strong>–</strong> b. v kolonii spoj. sliz. trubičkami, toxická, vod. květ<br />
Gomphosphaeria <strong>–</strong> slizovité kolonie, b. na obvodě, plankton<br />
rybníků, údolní nádrže<br />
Woronichinia naegeliana<br />
Gomphosphaeria<br />
G.<br />
G. semen-vitis
Pleurocapsa<br />
II. Pleurocapsales<br />
freahwater/marine, in alkalic waters calcification <strong>–</strong><br />
stromatolite-like structures, containes scytonemin<br />
(UV filter)<br />
Stanieria
III. Oscillatoriales <strong>–</strong> trichomy v pochvě<br />
Lyngbya <strong>–</strong> metafyton, sladk., hodně moř., povlaky, toxické,<br />
široká pochva (nad 1 μm), trichomy nad 8 μm
Oscillatoriales <strong>–</strong> trichomy v pochvě<br />
Microcoleus <strong>–</strong> svazky vláken s tenkými pochvami, málo pohyblivý,<br />
bez hormogonií, kalyptra, typický pro slané vody, v půdě M.<br />
vaginatus
Oscillatoriales <strong>–</strong> trichomy v pochvě<br />
Phormidium <strong>–</strong> tenké slizovité pochvy (nad 3 μm), ale ne u všech<br />
vláken, bentický, nárosty, půdní, mnoho druhů<br />
P. foveolarum<br />
P. tenue<br />
P. vulgare
Oscillatoriales <strong>–</strong> trichomy v pochvě<br />
Leptolyngbya <strong>–</strong> tenké slizovité pochvy, drobná (do 3 μm),<br />
nepohyblivá, ubikvitní (mnoho typů), bentická, nárosty, půdní<br />
L. fovearum<br />
L. africana<br />
L. fovearum<br />
L. fragilis<br />
L. tenuis<br />
L. tenuis
Oscillatoriales <strong>–</strong> neperzistující pochvy,<br />
b. delší než širší<br />
Limnothrix <strong>–</strong> jednotlivá vlákna v planktonu, nezaškrcovaná<br />
vlákna, nepatrný sliz, aerotopy na přepážkách<br />
Pseudanabaena, Dolichospermum <strong>–</strong> zaškrcovaná vlákna, plankton<br />
Limnothrix Pseudanabaena
Oscillatoriales <strong>–</strong> neperzistující pochvy,<br />
b. kratší než širší<br />
Oscillatoria <strong>–</strong> pohyblivá vlákna, pochvy za stresu, metafyton,<br />
nárosty, vod. květy<br />
O. limosa<br />
O. limosa
Oscillatoriales <strong>–</strong> neperzistující pochvy,<br />
b. kratší než širší<br />
Planktothrix <strong>–</strong> vlákna mírně pohyblivá, aerotopy, eutr. vody,<br />
plankton, vodní květ<br />
P. aghardii<br />
P. aghardii<br />
P. rubescens
Oscillatoriales <strong>–</strong> neperzistující pochvy,<br />
b. kratší než širší<br />
Trichodesmium <strong>–</strong> vlákna tvoří kulovité či svazečkovité kolonie<br />
dominantní pigment fykoerythrin, moř., tropické, subtropické obl.,<br />
vodní květ <strong>–</strong> největší „vazač“ N v otevřeném oceánu<br />
fluorescence<br />
pobřeží záp. Austrálie
Nostocales <strong>–</strong> interkalární heterocyty,<br />
Anabaena <strong>–</strong> druhově bohatý rod, bentické a planktonní (aerotopy),<br />
vodní květ, některé toxické, citlivost na nadměrné hnojení, nádrže<br />
A. planktonica<br />
A. spiroides<br />
A. flos-aqaue<br />
A. compacta<br />
A. affinis
Nostocales <strong>–</strong> interkalární heterocyty,<br />
Aphanizomenon <strong>–</strong> vlákna na koncích rovnoměrně zúžená, vodní<br />
květ „jehličí na hladině“ <strong>–</strong> A. flos-aque<br />
A. flos-aque<br />
A. gracile
Nostocales <strong>–</strong> interkalární heterocyty<br />
Nostoc <strong>–</strong> stočená nevětvená vlákna uložená ve slizu, akinety ze<br />
všech vegetativních buněk, sladkovodní, půdní, symbiont
Nostocales <strong>–</strong> interkalární heterocyty<br />
Nodularia <strong>–</strong> b. kratší než širší, akinety v řadě za sebou, bentické<br />
i planktonní, povlaky, sladk., moř., vodní květ, tox.<br />
N. spumigena<br />
Soos<br />
N. harveyana
Nostocales <strong>–</strong> terminální heterocyty<br />
Cylindrospermopsis <strong>–</strong> heterocyty kuželovité, asymetrické dělení<br />
konc. b., eutr. vody, mírné pásmo - tropy, vod. květ, toxický
Cylindrospermopsis raciborskii<br />
Invasion to temperate localities<br />
France 1994<br />
Bloom in the Northern Argentina
Nostocales <strong>–</strong> terminální heterocyty<br />
Cylindrospermum <strong>–</strong> tenká slizová pochva, heterocyty kulovité<br />
až oválné, litorál vod, vlhká půda<br />
klíčení akinet - hormogonia
Nostocales <strong>–</strong> bazální heterocyty<br />
Calothrix <strong>–</strong> vlákna jednotlivá nebo tvoří chomáče, slizové pochvy,<br />
ponořené substráty, půdní<br />
Dichothrix <strong>–</strong> svazky (trsy) vláken ve slizu, často žlutohnědé barvy<br />
Dichothrix<br />
C. parietina<br />
Calothrix
Nostocales <strong>–</strong> bazální heterocyty<br />
Gloeotrichia <strong>–</strong> kulovité kolonie, makroskopické, ponořené<br />
substráty, i v planktonu
Nostocales <strong>–</strong> basal heterocysts<br />
Rivularia <strong>–</strong> tuhé kulovité kolonie, makroskopické, ponořené<br />
substráty, kalcifikace krusty, travertiny<br />
Baltic sea-Hiddensee
Nostocales <strong>–</strong> false branching<br />
Tolypothrix <strong>–</strong> boční vlákna vyrůstají po jednom, podélně vrstevnatá<br />
pochva či bez pochvy, epifyt, kameny potoků, ponořené rostliny<br />
Hasallia <strong>–</strong> vzácná, aerofytická, chladící věže<br />
Tolypothrix Hasallia
Scytonema <strong>–</strong> slizová pochva, boční větévky po dvou, ojediněle<br />
po jednom, převážně sladkovodní, inkrustace CaCO 3 travertin<br />
Petalonema <strong>–</strong> šikmo vrstevnatá pochva, apikálně hormogonie<br />
Petalonema<br />
Nostocales <strong>–</strong> false branching<br />
S. crispata
Stigonematales <strong>–</strong> true branching<br />
Stigonema <strong>–</strong> výrazný sliz, vlákna uni či polyseriátní, větvení na<br />
všechny strany, subaericky, půda, nárosty, hl. tropy<br />
S. ocellatum S. ocellatum<br />
S. ocellatum
Hapalosiphon <strong>–</strong> uniseriátní, větvení jen na jednu stranu v pravém<br />
úhlu, metafyton, rašeliniště<br />
Fischerella - hlavní vlákno víceřadé, boční jednořadé, větvení<br />
jednostrannné, půda<br />
Hapalosiphon<br />
Stigonematales <strong>–</strong> true branching<br />
Hapalosiphon<br />
Fischerella<br />
F.<br />
Fischerella
Stigonematales <strong>–</strong> true branching<br />
Mastigocladus <strong>–</strong> vlákna uniseriátní, nepravidelně stočená, M.<br />
laminosus obligátní thermofil (teplota od 45 do 60°C, pH > 7,5,<br />
nízká salinita), 1862 Karlovy Vary
Ekology of <strong>Cyanobacteria</strong><br />
ubiquists, predominantly freshwaters, but also marine <strong>–</strong><br />
pikoplanktonic, extreme biotopes (glaciers, desert soil<br />
crusts, hot springs)<br />
colonization success they may survive in low light<br />
intensities in epilimnium, phenoplastic, produce toxins<br />
Why there are predominantly small species in the<br />
sea?
Eutrophication<br />
nutrient excess<br />
freshwater <strong>–</strong> P- limiting nutrient<br />
marine env. <strong>–</strong> N <strong>–</strong> limiting nutr.<br />
nutrient runoff from human<br />
activities:<br />
N <strong>–</strong> surface runoff, erosion<br />
P <strong>–</strong> industry, household
Eutrophication<br />
Czech republic 1985 - 2002
Water blooms
Cyanotoxins<br />
• alkaloid neurotoxins (block transmission<br />
of the signal from neuron to neurom<br />
(muscle)<br />
anatoxin, saxitoxin, aphanotoxin<br />
cylindrospermopsin<br />
microcystin, nodularin<br />
• alkaloid hepatotoxins<br />
• peptidic hepatotoxins<br />
• lipopolysaccharides<br />
component of cell walls in all cyanobacteria, alergenic
Cyanotoxins <strong>–</strong> over 50 species produced them<br />
Genus Produced toxins<br />
Anabaena anatoxins, microcystins, saxitoxins, LPS<br />
Aphanizomenon saxitoxins, cylindrospermopsins, LPS<br />
Cylindrospermopsis cylindrospermopsins, saxitoxins, LPS<br />
Hapalosiphon microcystins, LPS<br />
Lyngbya aplysiatoxins, Lyngbyatoxin-a, LPS<br />
Microcystis microcystins, LPS<br />
Nodularia nodularins, LPS<br />
Nostoc microcystins, LPS<br />
Phormidium (Oscillatoria) anatoxins, LPS<br />
Planktothrix (Oscillatoria)<br />
anatoxins, aplysiatoxins, microcystins,<br />
saxitoxins, LPS
• anti-herbivore chemicals<br />
• allelopatic interactions (affection of growth of other<br />
organisms)
Cyanotoxins<br />
Microcystis a Anabaena<br />
Microcystin<br />
peptidic hepatotoxin
Cyanotoxins<br />
peptidic hepatotoxin
Cyanotoxins<br />
Trichodesmium <strong>–</strong> Cook 200 years ago, extensive water bloos (10 <strong>–</strong><br />
100 km wide), saxitoxin, fish death<br />
alkaloid neurotoxin<br />
Pohled z družice
Epilitic cyanobacteria<br />
dark colour, UV protection<br />
Calothrix<br />
High scytonemin content<br />
(UV A protection) correlated with the<br />
magnitude
Epilitic cyanobacteria<br />
granite<br />
Quartz<br />
Chroococcidiopsis <strong>–</strong> underside of translucent stones
Epilitic cyanobacteria<br />
Chroococcidiopsis <strong>–</strong>able to survive extreme environments; NASA<br />
project <strong>–</strong> Mars colonization, E.I.Friedmann, astrobiologist<br />
lives in deserts under the<br />
translucent stones<br />
(humidity collectors, UV<br />
protection)<br />
Microbial ecology of absolute<br />
extreme environments<br />
Grilli-Caiola, M., D. Billi and E. I. Friedmann. 1996. Effect of desiccation on envelopes of the<br />
cyanobacterium Chroococcidiopsis sp. (Chroococcales). Eur. J. Phycol. 31:97-105.
<strong>Cyanobacteria</strong> in microbial crusts<br />
• bind particles <strong>–</strong> soil stabilisation prevent erosion<br />
• retain humidity, warming of the soil surface<br />
• nitrogen enrichment of the soil (10-25 kg/ha/year) nutrient<br />
imput
Soil crusts<br />
cyanobacteria, algae, mosses, lichens, fungi<br />
cyanobacteria and lichens dominant in hot deserts<br />
cyanobacteria to algae ratio changes with pH ( pH <strong>–</strong> green<br />
algae dominate)<br />
typical genera: Scytonema, Lyngbya, Oscillatoria, Microcoleus,<br />
Nostoc, Calothrix, Aphanocapsa, Chroococcidiopsis<br />
Microcoleus vaginatus
Termophilic cyanobacteria <strong>–</strong> hot springs<br />
extreme 70-73°C<br />
enzymes stable at higher temp.<br />
Synechococcus , Phormidium<br />
Octopus Spring, Yellowstone National<br />
Phormidium
M. laminosus is found in thermal areas<br />
throughout the world<br />
populations are typically genetically<br />
differentiated on local geographic scales<br />
suggests the existence of dispersal barriers<br />
conclusion corroborated by evidence for<br />
genetic isolation by distance<br />
a common ancestor associated with the<br />
western North American hot spot currently<br />
located below Yellowstone National Park<br />
Mastigocladus<br />
laminosus<br />
Miller et al. 2007, APPLIED<br />
AND ENVIRONMENTAL<br />
MICROBIOLOGY
Spirulina - Arthrospira<br />
tropical lakes, large-scale cultivations, dietary sypplements
history<br />
Spirulina - Arthospira<br />
Aztecs collected Spirulina<br />
already in the 16. century<br />
(Mexico)<br />
collection and drying of Spirulina<br />
near Chad Lake (Africa) - Dihé
Texcoco Lake, Mexico<br />
Spirulina - Arthrospira<br />
Earthrise Company, Hawaii USA<br />
• contains essential amino-acids<br />
• high protein and chlorophyll<br />
content<br />
• digestible cell wall<br />
• vitamins (B 12), carotenoids, Fe
origin of plastids<br />
Symbiotic interactions<br />
with algae <strong>–</strong> diatoms, dinophytes, Glaucophyta (cyanels)<br />
with fungi <strong>–</strong> lichens, Geosiphon<br />
with Bryophytes <strong>–</strong> Blasia, hornworts<br />
with ferns <strong>–</strong> Azolla<br />
with cycas<br />
Gunnera<br />
different degree of assimilation, it may be isolated into the<br />
culture
with algae <strong>–</strong> Glaucophyta (cyanels), diatoms<br />
Cyanophora<br />
paradoxa<br />
Richelia intracellularis<br />
Symbiotic interactions<br />
Glaucocystis<br />
nostochinearum
with fungi <strong>–</strong> lichens<br />
Symbiotic interactions<br />
Lobaria <strong>–</strong> cephalodium, Nostoc<br />
(sorál)<br />
Collema - Nostoc<br />
Peltula - Chroococcidiopsis
Symbiotic interactions<br />
with fungi<strong>–</strong> Geosiphon <strong>–</strong> Glomeromycota
Symbiotic interactions<br />
with Bryophytes <strong>–</strong> Blasia, hornworts Anthoceros<br />
Blasia Anthoceros<br />
liverworts and hornworts
Symbiotic interactions<br />
with ferns <strong>–</strong> Azolla has Nostoc in meristems<br />
biofertilizer in rice paddies, fixation 20 x higher within Azolla,<br />
Anabaena in Azolla has different surface antigens
with cycases (Nostoc)<br />
Cycas - roots<br />
Symbiotic interactions<br />
cortex of aboveground roots<br />
Salvinia
Symbiotic interactions<br />
Gunnera chilensis (mucillage-filled glands in the stems)- Nostoc<br />
heterocyty<br />
BS - Gunnera<br />
Stem
Prochloron and ascidians:<br />
Algae and symbioses str. 218
Prochloron and ascidians
Prochloron + Didemnum (Chordata, Tunicata)
Symbiotic interactions<br />
with fungi <strong>–</strong> Geosiphon <strong>–</strong> Glomeromycota
Why are cyanobacteria important?<br />
• the only one prokaryotes, that produce O 2 by water splitting<br />
in the light reaction of photo<strong>syn</strong>thesis<br />
• first terrestrial organisms<br />
• ability of cyanobacteria to perform oxygenic photo<strong>syn</strong>thesis<br />
is thought to have converted the early reducing atmosphere<br />
into an oxidizing one<br />
• cyanobacerial ancestor <strong>–</strong> actor of endosymbioses <br />
chloroplast of other algal groups (including vascullar plants)<br />
• the simplest organisms with circadian rythmus<br />
• favourite symbioses partners













