
City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus



1
Department of Geography, University College Cork, T12 K8AF Cork, Ireland
2
Environmental Research Institute, University College Cork, T23 XE10 Cork, Ireland
3
BirdWatch Ireland, A63 RW83 Kilcoole, Ireland
*
Author to whom correspondence should be addressed.
Geographies 2022, 2(2), 161-172; https://doi.org/10.3390/geographies2020011
Submission received: 16 February 2022
/
Revised: 12 March 2022
/
Accepted: 14 March 2022
/
Published: 23 March 2022
(This article belongs to the Special Issue Feature Papers of Geographies in 2022)
Abstract
:Herring gulls (Larus argentatus) are declining globally, but there are populations who are taking advantage of the new foraging and nesting opportunities afforded to them by urban landscapes. Nest-site selection (NSS) in urban environs is understudied, despite its critical role in supporting planning policy, biodiversity conservation and the management of human–wildlife conflict. The aim of this study was to assess the contribution of anthropogenic habitat features to NSS in urban populations of L. argentatus at different hierarchical levels in Fingal County, Ireland. We used generalised linear models with a logit function to investigate the relationship among nest sites, building features, street furniture (i.e., streetlights and refuse bins), landscape features, and presence of conspecifics at three different hierarchical levels, including the county, town, and colony levels. L. argentatus preferentially chose buildings that were closer to streetlights and food sources at the colony level, while avoiding streetlights when considered in isolation. Conspecific attraction at the county and colony levels indicated that individuals avoided neighbouring nest sites, yet this relationship was inverted at the town level, suggesting preference. Moreover, 75% of nests were within 30 m of each other (the average road width in the study area) when measured at the county level. Various relationships with different food sources were identified, suggesting within-population variation among preferences for nest sites. There appears to be a substantial population variation among preferences for nest sites, which does appear to be driven by the cross-scale decisions involved in nest-site selection.
1. Introduction
The world’s coastlines are urbanising rapidly, with an estimated one billion people expected to live within the lower-elevation coastal zone by 2060 [1]. As a result, natural coastal habitats and the species they support are increasingly under threat from urban expansion [2,3,4]. Coastal urban areas represent a mosaic of new habitats, and despite the pressures of urbanisation on numerous species, there are innovative populations of seabirds that are modifying their foraging and nesting behaviour to take advantage of these novel ecological niches [5,6,7]. Nest-site selection (NSS) is a subset of habitat selection that operates at fine spatial scales [8], allowing researchers and practitioners the opportunity to explore the landscape factors associated with where birds choose to nest. However, understanding species–habitat relationships in urban environments represents a unique yet understudied area of NSS research that is critical for planning policy, biodiversity conservation, and management of human–wildlife conflict [9,10].
Natural dwelling populations of herring gulls (Larus argentatus) are declining both globally [11] and locally [12]; however, they have been one of the most abundant urban nesting gulls in several countries [13]. For example, in Britain and Ireland, there have been over 11,047 pairs recorded nesting on buildings and other artificial structures since surveys began in 1969 [13,14]. Despite local urban successes, the habituation of roof-top nesting by L. argentatus has led to human–wildlife conflict. Given the red-listed conservation status of L. argentatus in several countries, it is crucial to understand the factors associated with NSS, in order to manage human–wildlife conflicts more effectively and support Sustainable Development Goals (SDGs) related to ecosystem health, such as SDG14 and SDG15 [15,16,17].
Studies have long investigated NSS in L. argentatus in the natural environment [18], with previous research addressing the landscape factors associated with NSS as the individual and combined effects of five main factors; the presence and behaviour of conspecifics, the availability of food, risk of predation, the availability of suitable nest substrate for raising offspring and the presence of a suitable ambient climate [19]. However, urban biotopes differ in many ways from natural biotopes [20] and in a recent review of avian NSS, Reynolds et al. [21] emphasised that assessments about where urban birds nest and the documentation of site availability in cities has been rather neglected as a research focus. Subsequently, there persists a need to investigate the availability and selection of artificial structures as nesting sites by L. argentatus [17], and whether equivalent urban landscape variables are important to urban NSS. Possible landscape features will be discussed in the context of urban NSS. Here, we omit ambient climate from discussion, as the factors we are interested in operate at a finer spatial resolution than climate which is often overarching in its effect of species distributions and habitat selection [22], although we do acknowledge that urban areas are warmer than natural sites, and thus breeding birds are less thermally challenged during incubation [23].
1.1. Presence of Conspecifics
Gulls are naturally semi-colonial nesters and studies suggest conspecific (individuals of the same species) presence may inadvertently provide social information to prospecting individuals regarding nest-site suitability [24,25]. For example, in a study attempting to characterise habitat preferences for 500 pairs of glaucous-winged gulls (Larus glaucescens) nesting on roofs along the Vancouver waterfront in Canada, Vermeer et al. [26] used a five-way contingency table to test the importance of building features (i.e., peaked or flat roofs, and length of structures), distance to water (i.e., near or far) and sociability (i.e., social index). They found that L. glaucescens nested significantly more frequently on roofs that were both close to the water and near other roofs with nesting gulls. The influence of conspecific attraction on NSS has been investigated in other coastal bird species, including Arctic-breeding shorebirds [27]. Considering the influence of social factors on NSS and the fact that L. argentatus are naturally semi-colonial nesters, NSS may occur near buildings already occupied by gulls.
1.2. Availabiltiy of Food
In addition to providing birds with nesting sites, urban environments also often provide a variety of food resources of human origin [16,28]. Diet analysis of Australian silver gulls (Larus novaehollandiae) revealed that 85% of stomach contents contained exclusively human refuse [29]. To understand this relationship, it is therefore necessary to determine how species use their environment concerning these food resources [28]. For example, Yoda et al. [7] found that nesting black-tailed gulls (Larus crassirostris) required shorter foraging trips (in terms of range and distance travelled) when exploiting urban feeding grounds in comparison with natural oceanic resources. Furthermore, research has shown that urban dwelling species, including house crows (Corvus splendens) and black kites (Milvus migrans) preferentially selected nest-sites nearer refuse bins and food centres or in areas under poor waste management regimes, respectively [30,31,32]. Considering the influence of human trash/waste as a food source is potentially advantageous to urban gulls in NSS.
1.3. Risk of Predation
In urban environments, ecological corridors provide safe foraging and resting opportunities for birds [33]. For example, the occurrence of urban black-headed gulls (Chroicocephalus ridibundus) was found to be positively related to the presence of watercourses and water reservoirs [33]. Studies have also shown that lesser black-backed gulls (Larus fuscus) can exist and breed successfully on a totally terrestrial based diet of earthworms from pastures [34], suggesting that open green spaces may also represent safe spaces and food sources for urban gulls. Recent research has shown that species richness and abundance of urban birds increases when there are connected green and blue spaces interspersed throughout the built-up areas [35]. Predation by great black-backed gulls (Larus marinus) and cannibalism has been known to reduce the productivity of L. argentatus colonies in natural environments [36]. A common feature represented throughout L. argentatus nest-sites on salt marsh islands in New Jersey was the trade-off between the need for concealment and the need for individuals to maintain some view of the surroundings of the nest [18]. NSS studies have found the influence of nest height and exposure as important variables for several other bird species [37,38,39]. Moreover, darkness is a rare occurrence in urban areas due to the presence of artificial light sources including streetlights. Studies have found diurnal feeding birds such as house sparrows (Passer domesticus) have shifted their feeding patterns and have been found to feed and nest by artificial light in urban environs [40,41,42]. As well as predation risk, there is also a mortality risk from proximity to heavy road traffic also found in urban areas [43]. It is unknown how L. argentatus responds to predation risk in urban environments, meaning there persists a need to investigate how artificial light and green spaces influence NSS.
1.4. Nest-Site Substrate
Nest sites require protection from extreme weather conditions [44]. For example, Kim and Monaghan [45] found that L. fuscus that nested adjacent to tall vegetation (and were subsequently sheltered from cold winds) raised chicks that grew faster than those on more exposed nests. In Britain, Monaghan [46] observed L. argentatus nesting amongst a variety of building structures sites and categorised these parameters into chimney stacks, sloping roofs, flat roofs and ledges. In Ireland, Roughan and O’Donovan [47] noted that nesting L. argentatus favoured flat roofs or built their nests against chimney stacks, walls, pipes and ventilation vents on shallow angle roofs. Building structure parameters such as nest height (i.e., roof height) and roof slope (i.e., roof type) are subsequently important habitat variables that contribute towards urban NSS of L. argentatus.
1.5. Research Questions
Several mechanisms have thus been invoked to explain NSS in a range of urban bird species, yet to date these have not reconciled the debate surrounding favourable conditions for NSS in urban L. argentatus. Moreover, given the conservation status and conflict narrative affiliated with L. argentatus, determining nest-site characteristics in urban dwelling populations may have important implications for developing effective habitat conservation and management strategies [8]. Moreover, habitat selection is a hierarchical process involving a series of innate and learned behavioural decisions made by an animal regarding habitat use at different scales of the environment [48,49]. Birds with a wide distribution, such as most seabirds, select their breeding habitats at several different levels, for example the colony itself, sub-area within a colony and then nest site [45]. Subsequently there remains a need to investigate the habitat features associated with NSS in urban settings across different hierarchical levels.
Here, we focus on the contribution of anthropogenic habitat features to NSS in urban populations of L. argentatus at different hierarchical levels in Fingal County, Ireland. We demonstrate that NSS is multifaceted and dependent on higher-order interactions and hierarchical scales. We highlight the importance of certain landscape features, including streetlights, food sources, and conspecific attraction at the county, town and colony levels. We acknowledge here that colony itself is a scale-dependent term, meaning colony could be used to describe the groupings at town, block, or street levels, but for our purposes, we use it to describe within-town clustering of nest sites, predominately at the block level. We discuss how there is now a need to incorporate social and behavioural factors to reflect the hierarchical nature of habitat selection and effectively support conservation efforts.
2. Materials and Methods
2.1. Study Area
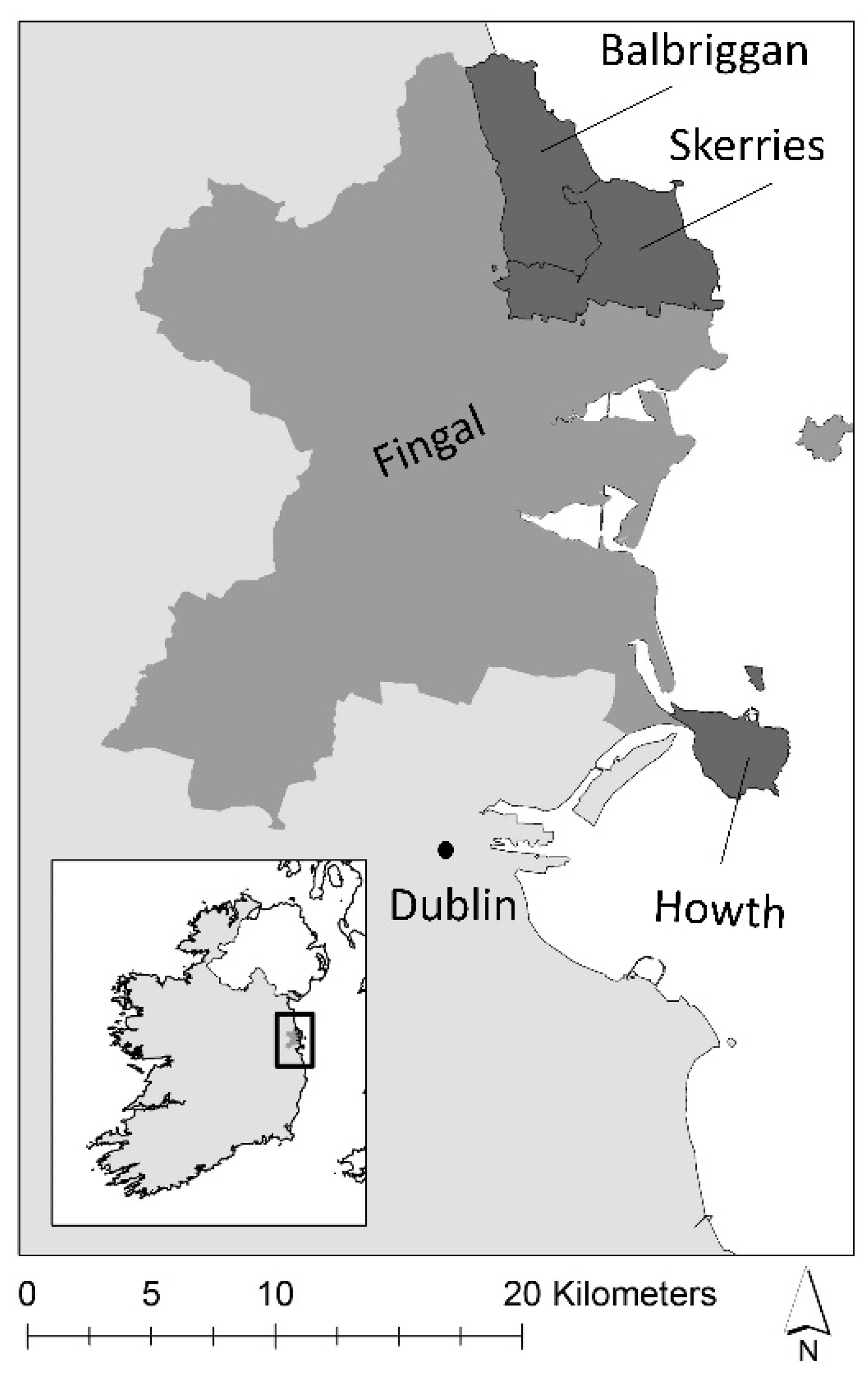
Herring gulls have been recorded extensively in three seaside towns in Fingal County, Ireland, north of Dublin (Figure 1). Howth, Skerries, and Balbriggan developed around natural harbours, and fishing and marine tourism have remained important industries to these towns. Skerries and Howth have been among Fingal’s longstanding major coastal residential towns, while Balbriggan has doubled in population during the last 20 years. With their coastline and countryside, these areas have rich biodiversity and there are several Special Areas of Conservation (SACs) and Special Protection Areas (SPAs) located around these urban centres. It is currently illegal to remove nests in Howth and Skerries, but a derogation is in place for this to be undertaken in Balbriggan under the National Wildlife Act. The Supplementary Material contains the lead author’s observations from walking surveys, which should provide further contextual information on the study areas.
2.2. Data Collection
A total of 260 buildings (Howth; 124, Balbriggan; 82 and Skerries: 54) used for nesting on by L. argentatus were obtained from the Roughan and O’Donovan (2018) roof nesting report. These locations were digitised by converting their geographic coordinates in Google Earth Pro (V 7.3.2.5776) and then corresponded to associated PRIME II building polygons (Ordinance Survey Ireland). The spatial distribution of utilised buildings in Balbriggan and Skerries was suggestive of three distinct colonies, while the spatial distribution of nested buildings in Howth was less expansive and appeared to be localised. This resulted in a total of 2109 buildings in the study area (699 in Howth, 840 in Balbriggan, and 570 in Skerries) where we had binary data representing nested or not nested.
We used the ESRI Collector for ArcGIS app on a Samsung Galaxy J3 android smartphone to document the presence and location of street furniture (i.e., streetlights and refuse bins), and inhibitors (i.e., barbed wire, metal spikes, and metal casings) in 2019. Building properties, including building height (number of floors) and roof type (gable, hip, flat and mansard) were recorded using street view imagery accessed through the Google Earth Pro (V 7.3.2.5776) platform. Distance to street furniture (i.e., streetlights and refuse bins), coastline, harbour, and open green spaces (>5000 m2—the equivalent size of a football pitch) were calculated for each building.
2.3. Data Analysis
Generalised linear models (GLMs) with a binomial distribution and logit link were used to explore the landscape features responsible for nest-site selection in L. argentatus at the county, town, and colony levels. GLMs with standardised coefficients were run in R 4.0.5 [50]. Final models were selected based on Akaike Information Criterion (AIC) using both forwards and backwards stepwise selection of models using the MASS package [51] and presented using the jtools package [52]. Third- and fourth-order interactions were investigated, but these did not improve final models. Therefore, final models include only main effects and second-order interactions. Due to a lack of variation in the building structures at the town and colony levels, height was removed from statistical models to allow for model convergence. Data and code are provided in the Supplementary Material. To ensure that model results were presented to allow robust interpretation, all models that scored an AIC value within Δi of less than 2 were explored and presented in the Supplementary Material.
3. Results
The spatial distribution of nested buildings in Balbriggan and Skerries was suggestive of three distinct colonies (Figure 2a,b), while the spatial distribution of nested buildings in Howth was less expansive (less < 0.5 km) and appeared to be localised (Figure 2c).
Our results indicated several important drivers of NSS at the county, town and colony levels. Multiple variables were returned in the final model selected by AIC when NSS was studied at the county level (Table 1), with NSS appearing to significantly favour buildings in a closer proximity to streetlights and harbours, a preference for hip roof structures over gable, while nesting further away from refuse bins, the coastline, and conspecifics. Several interactions at the county level were returned (Table 1), including several with distance to conspecifics (neighbours) and roof type, suggesting that NSS is multifaceted and dependent on higher-order interactions.
In general, the stepAIC process generated between 9 and 15 candidate models for all hierarchical scales, with the exception of colony 2 at Skerries which reported 27 candidate models. Alternative candidate models that had a Δi of less than 2 when AIC was considered are reported in the Supplementary Material. Across all scales, there were seldom more than three possible candidate models, with coefficient relationships largely similar. Additional variables that were added to models consisted of interactions, supporting our assertion that NSS is multifaceted and dependent on higher-order interactions. Moreover, there were only six instances where a coefficient inverted its relationship with habitat selection, out of a possible 164. Most inversions were identified in Skerries and mostly due to overfitting that occurred in that colony due lack of variability in specific urban landscape features (i.e., roof types).
At the town level, the species–habitat relationship did not remain consistent across all three towns (Figure 3, Table 1). NSS appeared to significantly favour proximity to refuse bins in Balbriggan while avoiding locations closer to such infrastructure in Howth and Skerries. Significant interactions between proximity to bins, the harbour, and the coastline were found in all towns, suggesting birds preferentially selected locations that were closer to multiple potential food sources. Proximity to streetlights was not significant as a main effect, although there were several significant interactions between streetlights and roof type, as well as proximity to bins, the harbour, and coastline, suggesting that NSS nearer food sources may be associated with streetlights. There was a preference for NSS closer to neighbours in Howth and Balbriggan, while avoidance in Skerries, although most interactions with distance to neighbour, which were primarily food sources, suggested avoidance.
The drivers of NSS were highly variable among colonies (Table 1). In Balbriggan, all three colonies appeared to favour proximity to the harbour, but proximity to other locations such as greenspaces, coastline, and bins varied among colonies. Proximity to streetlights, the coastline, and greenspaces were more routinely reported as significant variables in the model outputs, suggesting these variables play an important role in NSS at the finest scale of habitat selection. Significant negative coefficients for interactions with streetlights and food sources suggest that birds are preferentially choosing such locations. Again, presence of conspecifics and proximity to food sources resulted in positive coefficients, suggesting avoidance of both features for most colonies.
4. Discussion
The primary aims of this research were to assess the contribution of anthropogenic habitat features to NSS in rooftop nesting populations of L. argentatus in county Fingal, Ireland investigating the hierarchical impact of NSS. In total, 260 nests were identified out of 2109 buildings across the three towns in 2018. By comparing anthropogenic habitat features identified at presence and absence sites at multiple levels of analysis, several variables were deemed to be important drivers of NSS, with different interactions occurring at all levels of analysis (Table 1; Figure 3; Supplementary Material), suggesting that NSS is multifaceted and dependent on higher-order interactions.
The significant associations between streetlights, food sources and nest presence have provided evidence to support the suggestion that artificial light may play an important role in L. argentatus NSS (Table 1). L. argentatus preferentially chose buildings that were closer to streetlights at the county level, with several significant negative interactions among proximity to streetlights and food sources at the colony level, suggesting preferential selection. L. argentatus are diurnal in nature and naturally forage during daylight; however, artificial light at night may extend foraging opportunities for the urban populations [53]. Moreover, studies have shown that artificial structures (e.g., poles and pylons) act as perches and have become an important habitat feature for various bird species including corvids and raptors [54]. Birds make use of these artificial perches to hunt prey, rest between movements in the landscape, monitor territories and watch reproductive sites [55]. Subsequently, proximity to streetlights may provide L. argentatus the opportunity for parental vigilance to protect their offspring, corroborating research from natural environments [56]. The average streetlight is approximately the same height as a two-storey building and the lamp cover provides a level platform on which L. argentatus parents can perch, monitor the activity of their young and survey the nest-site area for predators. To develop a deeper understanding of the relationship between artificial light and NSS will require further observational studies at nest sites to assess activity levels of urban L. argentatus at night.
Different relationships with potential food sources were reported across hierarchical scales. For example, at the county level, distance to street bins exhibited a positive relationship with NSS, suggesting avoidance (Table 1; Figure 3). NSS in Balbriggan identified proximity to bins as a negative coefficient, although in Skerries and Howth nests were located further away from bins than unnested buildings (Table 1; Figure 3). Similar inversions in the NSS–environment relationships for distance to harbour, coastline, and greenspaces (all potential sources of food) were also observed, suggesting a large variation among NSS preferences related to food availability. L. argentatus forage on resources locally most available; therefore, spatial variation observed among colonies may arise due to the colonies having different local foraging opportunities [57]. Moreover, foraging distances of L. argentatus are scale dependent, with Van Donk et al. [58] identifying these differences in the order of less than 10 km at fine scales to hundreds of kilometres at macroscales, with recent foraging records observing movements generally within 40–60 km from nest sites [53,59]. Therefore, the variations observed within our results regarding the selection or avoidance of nest sites in relation to possible food sources may be compounded by further hierarchical scales operating within foraging processes. Similarly, recent research has identified that human handling of food may serve as a cue for L. argentatus [60], which could suggest that selection of foraging sites may be multifaceted in urban environs and would support the significant interactions found between distance to streetlights and potential food sources. Nonetheless, several significant interactions were observed between such parameters, suggesting future research is needed to parse out the fine-scale behavioural decisions within local populations of L. argentatus.
While urban gull’s habitat preferences differed from absence sites with respect to roof type, the preferred type was not the same in every town (Table 1; Figure 3). The nesting population in Balbriggan and Skerries appeared to prefer gable-style roofs, while nesting gulls preferentially selected both gable- and hip-style roofs in Howth, which suggests that while roof type may explain nest presence within towns, it may not be a transferable feature between towns. However, gable and hip are relatively coarse categories, and when studied at the colony level, little variation among types was present, meaning in some instances models may be over fitted. Therefore, to unravel the relationship between roof type and NSS, a more detailed assessment is needed that includes quantitative measurements of slope, roofing material, level of biodiversity on the roof (lichens), and degree of shelter and orientation [61].
At the county level, over three-quarters of the nest sites were located 30 m or less apart and tended to be positioned either on a roof next to a neighbour or across the street from a neighbour (Figure 4). In our study area, 30 m represents the average width of a road, and therefore considering the focal adjacency of a possible neighbour in all directions. However, conspecific attraction was positive at the county and colony levels, indicating that individuals favoured sites that did not have immediate neighbours, yet this relationship was negative in Howth and Balbriggan, suggesting preference (Table 1; Figure 3). The apparent dispersion at different hierarchical scales may indicate a new phenomenon of urban gulls in avoiding conspecifics, although alternatively (and perhaps more likely) it is due to human interference through the application of inhibitor structures, which may be forcing the nesting pairs further apart. During walking surveys of the area in 2019, we noted instances of repeat nesting on buildings that had inhibitors, albeit in low numbers. We decided against a direct inclusion of inhibitors in model parameterisation, due to the fact we were unsure whether inhibitor structures were present during the previous year of drone surveys; however, we posit a possible trend between inhibitors and nest dispersal.
Studies have shown that repeat nesting can lead to local knowledge of an area and have important implications for individual fitness through increased survival or reproduction [62]. Therefore, NSS at repeat nesting sites warrants further research, detailing the specific habitat features and monitoring the degree of reproductive success through the nesting season. However, roof nesting is often perceived as a serious nuisance in county Fingal (and beyond) and the opportunity for repeat nesting is dependent on the property owner. A more effective approach to studying repeat nesting in the future may involve building relationships with property owners that are indifferent or feel positively towards the gulls and are less likely to apply inhibitors. This is particularly pertinent given that recent research has identified that some urban Larus spp. use their nesting site all year around to support foraging during winter months [63]. Moreover, it has been identified that the relocation of nest sites in urban L. fuscus results in a reduced reproductive success, suggesting a strong philopatry of the species [64]. Collaborating with locals in this way may stimulate interest in their local wildlife and potentially provide an uninterrupted time series of repeat nesting data, with recent research identifying that developing empathy with a species or an individual animal increases the ethical duty of care, which subsequently supports biodiversity conservation in the long term [65].
5. Conclusions
Our results highlight the continued importance of physical factors in determining NSS for L. argentatus, but also highlight the need to incorporate social and behavioural factors into further studies. With the global decline of L. argentatus and the fact that NSS in urban environs is understudied, it is of critical importance that future research is directed towards understanding the hierarchical nature of habitat selection. We suggest that NSS is multifaceted and dependent on higher-order interactions, with gulls preferentially choosing buildings that were closer to streetlights and food sources at the colony level, while avoiding streetlights when considered in isolation. Conspecific attraction was positive at the county and colony levels, indicating that individuals avoided neighbouring nest sites, yet this relationship was negative at the town level, suggesting preference. There appears to be a substantial population variation among preferences for nest sites, which does appear to be driven by the multi-scale decisions involved in nest-site selection.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/geographies2020011/s1, Supplementary Material includes Data and Code, Field Site Observations, and All Candidate Logit Models.
Author Contributions
Conceptualisation, C.D.P., S.N., F.C. and P.H.; methodology, C.D.P. and P.H.; data curation, C.D.P.; formal analysis, C.D.P. and P.H.; writing—original draft preparation, C.D.P., S.N., F.C. and P.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
The datasets and code generated during this research are available in the Supplementary Material. The Prime 2 dataset which includes information on building polygons and greenspaces was sourced from Ordnance Survey Ireland (OSi) as part of the National Mapping Agreement, which are publicly available to Irish government bodies and public sector bodies. As part of this agreement, we do not have permission to share the data, but it can be sourced from https://www.osi.ie/about/forbairti-satodhchai/prime2/?lang=ga (accessed on 4 July 2019). Coastline data is available from https://www.eea.europa.eu/ (accessed on 5 July 2019).
Acknowledgments
We would like to thank the reviewers and editors for their suggestions and comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Neumann, B.; Vafeidis, A.T.; Zimmermann, J.; Nicholls, R.J. Future Coastal Population Growth and Exposure to Sea-Level Rise and Coastal Flooding—A Global Assessment. PLoS ONE 2015, 10, e0131375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennie, J.; Duffy, J.P.; Davies, T.W.; Correa-Cano, M.E.; Gaston, K.J. Global Trends in Exposure to Light Pollution in Natural Terrestrial Ecosystems. Remote Sens. 2015, 7, 2715–2730. [Google Scholar] [CrossRef] [Green Version]
- Hastings, R.; Cummins, V.; Holloway, P. Assessing the impact of physical and anthropogenic environmental factors in determining the habitat suitability of seagrass ecosystems. Sustainability 2020, 12, 8302. [Google Scholar] [CrossRef]
- Holloway, P.; Field, R. Can rock-rubble groynes support similar intertidal ecological communities to natural rocky shores? Land 2020, 9, 131. [Google Scholar] [CrossRef]
- Belant, J.L. Gulls in urban environments: Landscape-level management to reduce conflict. Landsc. Urban Plan. 1997, 38, 245–258. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Warren, P.S. Urban Bird Ecology and Conservation; Studies in Avian Biology; University of California Press: Berkeley, CA, USA, 2012; Volume 45. [Google Scholar]
- Yoda, K.; Tomita, N.; Mizutani, Y.; Narita, A.; Niizuma, Y. Spatio-temporal responses of black-tailed gulls to natural and anthropogenic food resources. Mar. Ecol. Prog. Ser. 2012, 466, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Jones, J. Habitat Selection Studies in Avian Ecology: A Critical Review. Ornithology 2001, 118, 557–562. [Google Scholar] [CrossRef]
- Clucas, B.; Marzluff, J.M. Attitudes and Actions toward Birds in Urban Areas: Human Cultural Differences Influence Bird Behavior. Auk 2012, 129, 8–16. [Google Scholar]
- McCaffrey, R.E.; Mannan, R.W. How scale influences birds’ responses to habitat features in urban residential areas. Landsc. Urban Plan. 2012, 105, 274–280. [Google Scholar] [CrossRef]
- BirdLife International. European Red List of Birds; Office for Official Publications of the European Communities: Luxembourg, 2015. [Google Scholar]
- Madden, B.; Newton, S. Herring gull Larus argentatus. In Seabird Population of Britain and Ireland; A & C Black: London, UK, 2004; pp. 242–261. [Google Scholar]
- Monaghan, P.; Coulson, J.C.C. Status of Large Gulls Nesting on Buildings. Bird Study 1977, 24, 89–104. [Google Scholar] [CrossRef]
- Raven, S.J.; Coulson, J.C. The distribution and abundance of Larus gulls nesting on buildings in Britain and Ireland. Bird Study 1997, 44, 13–34. [Google Scholar] [CrossRef]
- Soldatini, C.; Albores-Barajas, Y.V.; Mainardi, D.; Monaghan, P. Roof nesting by gulls for better or worse? Ital. J. Zool. 2008, 75, 295–303. [Google Scholar] [CrossRef]
- Mainwaring, M.C. The use of man-made structures as nesting sites by birds: A review of the costs and benefits. J. Nat. Conserv. 2015, 25, 17–22. [Google Scholar] [CrossRef]
- Beasley, E.R. Foraging Habits, Population Changes, and Gull-Human Interactions in an Urban Population of Herring Gulls (Larus argentatus) and Lesser Black-backed (Larus fuscus). Master’s Thesis, Middlesex University, London, UK, 2017. [Google Scholar]
- Burger, J.; Shisler, J. Nest Site Selection and Competitive Interactions of Herring and Laughing Gulls in New Jersey. Auk 1978, 95, 252–266. [Google Scholar]
- Mainwaring, M.C.; Hartley, I.R.; Lambrechts, M.M.; Deeming, D.C. The design and function of birds’ nests. Ecol. Evol. 2014, 4, 3909–3928. [Google Scholar] [CrossRef]
- Isaac, B.; Cooke, R.; Simmons, D.; Hogan, F. Predictive mapping of powerful owl (Ninox strenua) breeding sites using Geographical Information Systems (GIS) in urban Melbourne, Australia. Landsc. Urban Plan. 2008, 84, 212–218. [Google Scholar] [CrossRef]
- Reynolds, S.J.; Ibáñez-Álamo, J.D.; Sumasgutner, P.; Mainwaring, M.C. Urbanisation and nest building in birds: A review of threats and opportunities. J. Ornithol. 2019, 160, 841–860. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.A.; Holloway, P. Incorporating movement in species distribution models. Prog. Phys. Geogr. 2015, 39, 837–849. [Google Scholar] [CrossRef]
- Heppner, J.J.; Ouyang, J.Q. Incubation behavior differences in urban and rural house wrens, Troglodytes aedon. Front. Ecol. Evol. 2021, 9, 89. [Google Scholar] [CrossRef]
- Thériault, S.; Villard, M.A.; Haché, S. Habitat selection in site-faithful ovenbirds and recruits in the absence of experimental attraction. Behav. Ecol. 2012, 23, 1289–1295. [Google Scholar] [CrossRef] [Green Version]
- Mariette, M.M.; Griffith, S.C. Conspecific attraction and nest site selection in a nomadic species, the Zebra finch. Oikos 2012, 121, 823–834. [Google Scholar] [CrossRef]
- Vermeer, K.; Power, D.; Smith, G.E.J. Habitat Selection and Nesting Biology of Roof-Nesting Glaucous-Winged Gulls. Colon. Waterbirds 1988, 11, 189–201. [Google Scholar] [CrossRef]
- Cunningham, J.A.; Kesler, D.C.; Lanctot, R.B. Habitat and social factors influence nest-site selection in Arctic-breeding shorebirds. Auk 2016, 133, 364–377. [Google Scholar] [CrossRef]
- O’Hanlon, N.J. Spatial Variation in Herring Gull Traits and Their Potential as Monitors of the Coastal Environment. Ph.D. Thesis, University of Glasgow, Glasgow, UK, 2016. [Google Scholar]
- Coogan, S.C.P.; Raubenheimer, D.; Zantis, S.P.; Machovsky-Capuska, G.E. Multidimensional nutritional ecology and urban birds. Ecosphere 2018, 9, e02177. [Google Scholar] [CrossRef] [Green Version]
- Soh, M.C.K.; Sodhi, N.S.; Seoh, R.K.H.; Brook, B.W. Nest site selection of the house crow (Corvus splendens), an urban invasive bird species in Singapore and implications for its management. Landsc. Urban Plan. 2002, 59, 217–226. [Google Scholar] [CrossRef]
- Kumar, N.; Gupta, U.; Jhala, Y.V.; Qureshi, Q.; Gosler, A.G.; Sergio, F. Habitat selection by an avian top predator in the tropical megacity of Delhi: Human activities and socio-religious practices as prey-facilitating tools. Urban Ecosyst. 2018, 21, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Spelt, A.; Williamson, C.; Shamoun-Baranes, J.; Shepard, E.; Rock, P.; Windsor, S. Habitat use of urban-nesting lesser black-backed gulls during the breeding season. Sci. Rep. 2019, 9, 10527. [Google Scholar] [CrossRef]
- Maciusik, B.; Lenda, M.; Skórka, P. Corridors, local food resources, and climatic conditions affect the utilization of the urban environment by the Black-headed Gull Larus ridibundus in winter. Ecol. Res. 2010, 25, 263–272. [Google Scholar] [CrossRef]
- Garthe, S.; Schwemmer, P.; Paiva, V.H.; Corman, A.M.; Fock, H.O.; Voigt, C.C.; Adler, S. Terrestrial and Marine Foraging Strategies of an Opportunistic Seabird Species Breeding in the Wadden Sea. PLoS ONE 2016, 11, e0159630. [Google Scholar] [CrossRef] [Green Version]
- Lambert, L.; Cawkwell, F.; Holloway, P. The Importance of Connected and Interspersed Urban Green and Blue Space for Biodiversity: A Case Study in Cork City, Ireland. Geographies 2021, 1, 217–237. [Google Scholar] [CrossRef]
- Parsons, J. Cannibalism in Herring Gulls. Br. Birds 1971, 64, 524–537. [Google Scholar]
- Bergin, T.M. Habitat Selection by the Western Kingbird in Western Nebraska: A Hierarchical Analysis. Condor 1992, 94, 903–911. [Google Scholar] [CrossRef]
- Mateo-Tomás, P.; Olea, P.P. Combining scales in habitat models to improve conservation planning in an endangered vulture. Acta Oecol. 2009, 35, 489–498. [Google Scholar] [CrossRef]
- Fischer, J.H.; Debski, I.; Taylor, G.A.; Wittmer, H.U. Nest site selection of South Georgia Diving-petrels Pelecanoides georgicus on Codfish Island, New Zealand: Implications for conservation management. Bird Conserv. Int. 2017, 28, 216–227. [Google Scholar] [CrossRef]
- Bakken, L.E.; Bakken, G.S. American Redstart Feeding by Artificial Light. Ornithology 1977, 94, 373–374. [Google Scholar]
- Sohi, G.K.; Kler, T.K. Adaptations in avian nesting behavior in relation to indigenous trees and housing structures in Punjab. J. Entomol. Zool. Stud. 2017, 5, 1045–1051. [Google Scholar]
- Holveck, M.J.; Grégoire, A.; Doutrelant, C.; Lambrechts, M.M. Nest height is affected by lamppost lighting proximity in addition to nestbox size in urban great tits. J. Avian Biol. 2018, e01798. [Google Scholar] [CrossRef] [Green Version]
- Ben-Aharon, N.; Kapota, D.; Saltz, D. Roads and road-posts as an ecological trap for cavity nesting desert birds. Front. Conserv. Sci. 2020, 1, 614899. [Google Scholar] [CrossRef]
- Yorio, P.; Bertellotti, M.; Quintana, F. Preference for Covered Nest Sites and Breeding Success in Kelp Gulls Larus dominicanus. Mar. Ornithol. 1995, 23, 121–128. [Google Scholar]
- Kim, S.Y.; Monaghan, P. Effects of vegetation on nest microclimate and breeding performance of lesser black-backed gulls (Larus fuscus). J. Ornithol. 2005, 146, 176–183. [Google Scholar] [CrossRef]
- Monaghan, P. Aspects of the breeding biology of Herring Gulls Larus argentatus in urban colonies. Ibis 1979, 121, 475–481. [Google Scholar] [CrossRef]
- Roughan and O’Donovan Consulting Engineers. Nesting Gulls Populations in Balbriggan; Skerries and Howth, Co. Dublin Final Report to Fingal County Council: Dublin, Ireland, 2018. [Google Scholar]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Hutto, R.L. Habitat selection by nonbreeding migratory land birds. In Habitat Selection in Birds; Harcourt Brace Jovanovich: New York, NY, USA, 1985; pp. 455–476. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Long, J.A. jtools: Analysis and Presentation of Social Scientific Data. R Package Version 2.1.0. 2020. Available online: https://cran.r-project.org/package=jtools (accessed on 1 June 2021).
- Enners, L.; Schwemmer, P.; Corman, A.M.; Voigt, C.C.; Garthe, S. Intercolony variations in movement patterns and foraging behaviors among herring gulls (Larus argentatus) breeding in the eastern Wadden Sea. Ecol. Evol. 2018, 8, 7529–7542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tryjanowski, P.; Sparks, T.H.; Jerzak, L.; Rosin, Z.M.; Skórka, P. A Paradox for Conservation: Electricity Pylons May Benefit Avian Diversity in Intensive Farmland. Conserv. Lett. 2013, 7, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.F.; McCarron, V.E.A.; Zocche, J.J. Use of artificial perches by birds in ecological restoration areas of the Cerrado and Atlantic Forest biomes in Brazil. Neotrop. Biol. Conserv. 2018, 13, 24–36. [Google Scholar]
- Calladine, J. A comparison of Herring Gull Larus argentatus and Lesser Black-backed Gull Larus fuscus nest sites: Their characteristics and relationships with breeding success. Bird Study 1997, 44, 318–326. [Google Scholar] [CrossRef]
- O’Hanlon, N.J.; Nager, R.G. What Herring Gulls Eat and How Diet Affects Their Breeding Performance. In Encyclopedia of Animal Behavior, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Van Donk, S.; Shamoun-Baranes, J.; Bouten, W.; Van Der Meer, J.; Camphuysen, K.C. Individual differences in foraging site fidelity are not related to time-activity budgets in Herring Gulls. Ibis 2019, 162, 429–445. [Google Scholar] [CrossRef]
- O’Hanlon, N.J.; Nager, R.G. Identifying habitat-driven spatial variation in colony size of Herring Gulls Larus argentatus. Bird Study 2018, 65, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Goumas, M.; Boogert, N.J.; Kelley, L.A. Urban herring gulls use human behavioural cues to locate food. R. Soc. Open Sci. 2020, 7, 191959. [Google Scholar] [CrossRef] [Green Version]
- Rock, P. Urban gulls: Problems and solutions. Br. Birds 2005, 98, 338–355. [Google Scholar]
- Clark, D.E. Roosting, Site Fidelity, and Food Sources of Urban Gulls in Massachusetts: Implications for Protecting Public Water Supplies. Ph.D. Thesis, University of Massachusetts, Amherst, MA, USA, 2014. [Google Scholar]
- Pais de Faria, J.; Paiva, V.H.; Veríssimo, S.; Gonçalves, A.M.; Ramos, J.A. Seasonal variation in habitat use, daily routines and interactions with humans by urban-dwelling gulls. Urban Ecosyst. 2021, 24, 1101–1115. [Google Scholar] [CrossRef]
- Salas, R.; Muller, W.; Vercuijsse, H.; Lens, L.; Steinen, L. Forced nest site relocations negatively affect reproductive investment in a colonial seabird species. Biol. Conserv. 2020, 246, 108550. [Google Scholar] [CrossRef]
- Doubleday, K. Nonlinear liminality: Human-animal relations on preserving the world’s most famous tigress. Geoforum 2017, 81, 32–44. [Google Scholar] [CrossRef]
Figure 1.
The study area in Final County, Ireland, with the towns Balbriggan, Skerries, and Howth situated along the east coast of Ireland.
Figure 1.
The study area in Final County, Ireland, with the towns Balbriggan, Skerries, and Howth situated along the east coast of Ireland.

Figure 2.
Heat maps of nested sites in (a) Balbriggan, (b) Skerries, and (c) Howth.

Figure 3.
Coefficient plot of the county- and town-level models.

Figure 4.
At the county level, the proportion of nests in 2018 located within a distance range from a neighbouring conspecific nest.
Figure 4.
At the county level, the proportion of nests in 2018 located within a distance range from a neighbouring conspecific nest.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Regression outputs for the main effects and second-order interactions at the county, town, and colony levels. ** significant at α < 0.01; * significant at α < 0.05.
Table 1.
Regression outputs for the main effects and second-order interactions at the county, town, and colony levels. ** significant at α < 0.01; * significant at α < 0.05.
| County | Howth | Balbriggan | Skerries | Bal 1 | Bal 2 | Bal 3 | Ske 1 | Ske 2 | Ske 3 | |
|---|---|---|---|---|---|---|---|---|---|---|
| (Intercept) | −2.69 ** | −1.95 ** | −2.20 ** | −1.37 | 140.89 ** | −35.89 ** | −9.27 ** | 193.80 ** | 599.17 | −28,730.00 |
| Roof | 1.49 ** | 1.49 ** | −9.67 * | −0.09 | −55.15 | 3.99 ** | −574.53 ** | 28,740.00 | ||
| Neighbour | 0.53 ** | −1.46 ** | −0.01 | 1.47 ** | −99.43 ** | 0.95 ** | 4.69 | 1.54 * | 361.26 | 1.07 * |
| Bin | 0.31 ** | 7.07 ** | −1.21 * | 2.18 ** | −49.88 ** | −7.18 * | 28.58 ** | 166.52 ** | −25,080.00 | |
| Streetlight | −0.17 ** | 0.19 | −0.14 | 0.46 | 55.65 ** | −17.28 ** | −0.39 | 1.54 * | 2472.00 | |
| Harbour | −1.39 ** | −0.52 | 2.10 * | −0.84 | −141.49 ** | −39.25 ** | −50.28 ** | −560.12 ** | −5.46 | |
| Coastline | 0.36 | 0.65 | −2.96 ** | 0.01 | −80.10 ** | 32.77 ** | 36.70 ** | 193.96 ** | 3.43 | |
| Greenspace | 0.01 ** | 0.00 | 0.00 | −0.03 ** | 2.03 ** | 0.22 * | −0.20 ** | −2.78 ** | −38.69 | −0.18 ** |
| Roof*Bin | −0.51 * | −3.78 ** | −536.19 ** | 25,080.00 | ||||||
| Roof*Harbour | 0.88 ** | −14.28 ** | −165.90 | 1665.78 ** | ||||||
| Roof*Greenspace | −0.01 ** | −0.01 ** | −0.04 ** | −0.05 ** | ||||||
| Roof*Neighbour | 2.57 ** | 2.01 ** | 9.04 ** | |||||||
| Roof*Streetlight | −1.23 ** | 0.94 * | 6.89 ** | −2479.00 | ||||||
| Roof*Coastline | 2.15 ** | 11.97 ** | −2.88 ** | 112.93 | −430.79 ** | |||||
| Neighbour*Bin | 0.38 ** | 0.77 ** | 0.58 ** | 37.29 ** | 10.05 * | |||||
| Neighbour*Harbour | 0.51 ** | −0.39* | 19.66 ** | |||||||
| Neighbour*Coastline | −0.23 ** | 0.53 ** | 55.70 ** | 14.36 * | ||||||
| Neighbour*Greenspace | −0.01 ** | −0.11 ** | −0.09 ** | −0.11 ** | ||||||
| Neighbour*Streetlight | 0.40 | 4.66 * | −2.70 ** | 7.75 ** | ||||||
| Bin*Harbour | −0.31 | 6.89 | −0.98 ** | −3.47 ** | ||||||
| Bin*Coastline | −0.68 ** | −6.81 ** | 12.30 ** | |||||||
| Bin*Streetlight | −0.22 * | −32.45 ** | −5.04 ** | −7.24 ** | ||||||
| Bin*Greenspace | 0.02 ** | 0.02 ** | −0.57 ** | 0.07 * | 0.08 | −2.42 ** | -0.06 | |||
| Streetlight*Harbour | −0.32 ** | −0.96 ** | −50.99 ** | −19.71 ** | ||||||
| Streetlight*Coastline | −0.40 ** | 12.63 ** | −6.33 ** | −8.14 ** | ||||||
| Streetlight*Greenspace | 0.02 * | 0.04 ** | −0.02 * | |||||||
| Harbour*Coastline | −0.20 ** | 1.67 ** | 2.20 * | 59.45 ** | ||||||
| Harbour*Greenspace | 0.00 ** | −0.04 ** | 0.02 ** | 0.24 * | 0.34 ** | 8.17 ** | 0.06 | |||
| Coastline*Greenspace | 0.00 * | 0.03 ** | −0.82 ** | −0.21 * | −2.76 ** | −0.08 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dalla Pria, C.; Cawkwell, F.; Newton, S.; Holloway, P. City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus. Geographies 2022, 2, 161-172. https://doi.org/10.3390/geographies2020011
AMA Style
Dalla Pria C, Cawkwell F, Newton S, Holloway P. City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus. Geographies. 2022; 2(2):161-172. https://doi.org/10.3390/geographies2020011
Chicago/Turabian StyleDalla Pria, Caitlin, Fiona Cawkwell, Stephen Newton, and Paul Holloway. 2022. "City Living: Nest-Site Selection Preferences in Urban Herring Gulls, Larus argentatus" Geographies 2, no. 2: 161-172. https://doi.org/10.3390/geographies2020011