
A New Species of Toad (Anura: Bufonidae: Rhinella) from Northern Peru
by
 , , and
, , and
Edgar Lehr
1,2,* ,
,
Juan C. Cusi
2,
Lily O. Rodriguez
2,3,
Pablo J. Venegas
4,5,
Luis A. García-Ayachi
5,6 and
and
Alessandro Catenazzi
5,6,7 1
Department of Biology, Illinois Wesleyan University, 303 E Emerson, Bloomington, IL 61701, USA
2
Departamento de Herpetología, Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Avenue Arenales 1256, Lince, Lima 14, Peru
3
Centro de Conservación, Investigación y Manejo de Áreas Naturales, Avenue Benavides N° 1238, Lima 18, Peru
4
Rainforest Partnership, 4005 Guadalupe Street, Austin, TX 78751, USA
5
Instituto Peruano de Herpetología (IPH), Augusto Salazar Bondy 136, Urban Higuereta, Surco, Lima 33, Peru
6
División de Herpetología, Centro de Ornitología y Biodiversidad (CORBIDI), Santa Rita No. 105 Oficina 202, Urban Huertos de San Antonio, Surco, Lima 33, Peru
7
Department of Biological Sciences, Florida International University, 11200 SW 8th Street, Miami, FL 33199, USA
*
Author to whom correspondence should be addressed.
Taxonomy 2021, 1(3), 210-225; https://doi.org/10.3390/taxonomy1030015
Submission received: 22 June 2021
/
Revised: 26 July 2021
/
Accepted: 28 July 2021
/
Published: 31 July 2021
Abstract
:We describe a new species of Rhinella from montane forests between 1788 and 2305 m a.s.l. in the Departamentos Amazonas and San Martín, Peru. We tentatively assign the new species to the Rhinella festae species Group based on morphological similarities with its other 19 members. It is characterised by large size (maximum SVL 91.6 mm in females), a pointed and protruding snout that is posteroventrally inclined, absence of a visible tympanic annulus and tympanic membrane, long parotoid glands in contact with upper eyelid, presence of a dorsolateral row of enlarged tubercles, outer dorsolateral tarsus surface with a subconical ridge of fused tubercles, and absence of subgular vocal sac and vocal slits in males. One specimen from Departamento Amazonas tested positive for Batrachochytrium dendrobatidis.
1. Introduction
The Neotropical genus Rhinella Fitzinger, 1826 [1] contains 97 species of toads [2]. Pereyra et al. [3] performed a total-evidence-based phylogenetic analysis of Rhinella that included revised molecular and phenotypic datasets for 83 nominal species. The authors recognized the monophyly of Rhinella (once R. ceratophrys is excluded) and two major clades (Rhinella marina Clade, Rhinella margaritifera Clade) that contain eight Rhinella species groups [3].
Twenty-nine species of Rhinella occur in Peru, of which 13 (= 45%) are endemic to Peru [2]. The 29 Peruvian species of Rhinella are assigned to five species groups: festae Group, margaritifera Group, marina Group, spinulosa Group, and veraguensis Group [3,4,5,6]. The festae species Group was recognized by Moravec et al. [6] based on molecular characters and included eight species [6,7]: R. chavin (Lehr, Köhler and Ponce, 2001) [8]; R. festae (Peracca, 1904) [9]; R. lilyrodriguezae Cusi, Moravec, Lehr and Gvoždík, 2017 [7]; R. macrorhina (Trueb, 1971) [10], R. manu Chaparro, Pramuk and Gluesenkamp, 2007 [5], R. nesiotes (Duellman and Toft, 1979) [11], R. rostrata (Noble, 1920) [12], and R. yanachaga Lehr, Pramuk, Hedges and Córdova, 2007 [13]. Pereyra et al. [3] revised the festae species Group, provided a diagnosis for its members, and recognized anatomical and morphological synapomorphies. Thus, the festae species Group is currently considered to contain the following 19 species [3,6,7,14] which are distributed in Panama, Colombia, Ecuador, Peru, and Bolivia (Figure 1): R. acrolopha (Trueb, 1971) [10], R. arborescandens (Duellman and Schulte, 1992) [4], R. chavin, R. chullachaki Castillo-Urbina, Glaw, Aguilar-Puntriano, Vences and Köhler, 2021 [14], R. festae, R. lilyrodriguezae, R. lindae (Rivero and Castaño, 1990) [15], R. macrorhina, R. manu, R. multiverrucosa (Lehr, Pramuk and Lundberg, 2005) [16], R. nesiotes, R. nicefori (Cochran and Goin, 1970) [17], R. paraguas Grant and Bolívar-G., 2014 [18], R. rostrata, R ruizi (Grant, 2000) [19], R. tacana (Padial, Reichle, McDiarmid and De la Riva, 2006) [20], R. tenrec (Lynch and Renjifo, 1990) [21], R. truebae (Lynch and Renjifo, 1990) [21], and R. yanachaga.
Herpetological surveys of the Río Abiseo National Park in northern Peru, Departamento San Martín, in the late 1990s led to the discovery of six new species of anurans: four Phrynopus [22], one Pristimantis [23], one Gastrotheca [24], and an unknown species of Rhinella. Recent herpetological surveys in northern Peru (Departamentos San Martín and Amazonas) by PJV and LAGA led to the discovery of several specimens of the same unknown species of Rhinella. Herein we describe this new species of Rhinella from northern Peru (Figure 1 and Figure 2), which we tentatively assign to the festae species Group based on morphological characters.
2. Materials and Methods
The IACUC Committee of Illinois Wesleyan University approved this study (Protocol 19-011) on 5 November 2019. Specimens were fixed in 10% formol and stored in 70% ethanol. Measurements are given in millimeters (mm) and were taken by EL to the nearest 0.1 mm using a digital caliper. Notes on color in life were taken from field notes and color photographs. Webbing formula follows the standards of Myers and Duellman [25], whereas all other terminology is that of Duellman [26] and Duellman and Lehr [27]. Taxonomy of Rhinella and its species groups follow Pereyra et al. [3]. Format of the description follows the standards of Duellman and Mendelson [28] and Lötters and Köhler [29]. Measurement abbreviations used throughout the text are SVL, snout–vent length; TL, tibia length, FL, foot length (distance from proximal margin of inner metatarsal tubercle to tip of Toe IV); HL, head length (from angle of jaw to tip of snout); HW, head width (at level of angle of jaw); ED, horizontal eye diameter; IOD, interorbital distance; EW, upper eyelid width; IND, internarial distance; E–N, eye-nostril distance (straight line distance between anterior corner of orbit and posterior margin of external nares); PL, parotoid length; PW, greatest parotoid width. Fingers and toes are numbered preaxially to postaxially from I to IV, respectively I–V. We determined comparative lengths of Toes III and V by adpressing both toes against Toe IV; lengths of Fingers I and II were determined by adpressing the fingers against each other. Condition of the tympanum was assessed by visual examination under a microscope. Specimens were sexed externally based on the presence or absence of nuptial pads and internally based on the type of gonads. We attempted to extract DNA from tongue tissues of both the holotype and one paratype (MUSM 15958), which had been fixed using formalin for several days, with a Macherey–Nagel DNA FFPE (formalin-fixed paraffin-embedded) extraction kit. No tissue had been preserved in ethanol at the time of collection. Unfortunately, the attempted extraction was unsuccessful and we could not amplify fragments of 16S rRNA (the gene fragment that typically amplifies best in amphibians). Tissues from the more recently collected specimens are not available for this study. We swabbed one juvenile (CORBIDI 20748) from Nueva Esperanza, Bagua for infection by the pathogenic fungus Batrachochytrium dendrobatidis (Bd). We stroked the skin of the toad 30 times (5 strokes on each side of the abdominal midline, 5 strokes on the inner thighs of each hind leg, and 5 strokes on the foot webbing of each hind leg) with a synthetic dry swab (Medical Wire & Equipment, #113). We extracted DNA from the swab with PrepMan Ultra. We then analyzed the diluted DNA extract in a Life Technologies QuantStudio 3 qPCR instrument following the protocols of Hyatt et al. [30] and Boyle et al. [31]. We reported ZE, the genomic equivalent for Bd zoospores, after comparing the qPCR results to a set of standards. Museum acronyms are American Museum of Natural History (AMNH), New York, USA; Centro de Ornitología y Biodiversidad (CORBIDI), Lima, Peru; Museo de Historia Natural, Universidad Nacional Mayor de San Marcos (MUSM), Lima, Peru; and The Field Museum Chicago (FMNH), Chicago, USA. For specimens examined, see Appendix A. The extent of occurrence (EOO) was calculated using the webserver GeoCat (http://geocat.kew.org/) and the area of occupancy (AOO) was calculated based on cell width of 2 × 2 km.
3. Results
Systematics
Family Bufonidae Gray, 1825
Genus Rhinella Fitzinger, 1826
Rhinella moralesi sp. nov. Lehr, Cusi, Rodrigues, Venegas, García-Ayachi and Catenazzi, 2021
LSID:urn:lsid:zoobank.org:pub:BE76DE75-AF3A-4B99-9351-B11A65D0AC29
Holotype (Figure 3A and Figure 4A,B): MUSM 15959 (field number AMNH 11993), an adult male, collected in the Río Abiseo Valley, collected within a radius of 1 km around Jucusbamba (7°53′17.002″ S, 77°11′49.009″ W, WGS84 GPS coordinates, not to be confused with Jucusbamba in Departamento La Libertad) at ca. 1830 m a.s.l., Río Abiseo National Park, Departamento San Martín, Peru, on 11 July 1999 by L.O. Rodriguez and A. Catenazzi.
Paratypes (3 males, 3 females, Figure 4C,D, Figure 6 and Figure 7): MUSM 15958 (field number AMNH 11992), adult male, collected in the Río Abiseo Valley, near Jucusbamba (7°53′17.002″ S, 77°11′49.009″ W, WGS84 GPS coordinates) at 2010 m a.s.l., Río Abiseo National Park, Departamento San Martín, Peru, on 11 July 1999 by L.O. Rodriguez and A. Catenazzi. CORBIDI 713, adult female, collected in Laguna Negra (6°53′29.303″ S, 77°23′18.29″ W, WGS84 GPS coordinates) at 1788 m a.s.l., Province Mariscal Cáceres, Departmento San Martín, Peru, on 6 February 2008 by P.J. Venegas. CORBIDI 20268, male, collected in Posic (6°26′58.031″ S, 77°14′11.349″ W, WGS84 GPS coordinates) at 1902 m a.s.l., Province Mariscal Cáceres, Departmento San Martín, Peru, on 7 December 2018 by P.J. Venegas. CORBIDI 20371, adult male, CORBIDI 20372, adult female, collected in Nuevo Chirimoto (6°34′5.716″ S, 77°15′26.49″ W, WGS84 GPS coordinates) at 2019 m a.s.l., Province Mariscal Cáceres, Departmento San Martín, Peru, on 10 December 2018 by L.A. García-Ayachi. CORBIDI 20837, a subadult female, collected in Uriarte (5°29′52.389″ S, 78°21′30.799″ W, WGS84 GPS coordinates) at 2305 m a.s.l., Province Bagua, Departmento Amazonas, Peru, on 15 August 2019 by A. Marchelie.
Referred specimens (12 juveniles, Figure 6C,D,G,H): CORBIDI 18862, collected near Yambrasbamba (5°41′54.514″ S, 77°59′15.355″ W, WGS84 GPS coordinates) at 2072 m a.s.l., Province Bongara, Departmento Amazonas, Peru, 5 September 2017 by P.J. Venegas. CORBIDI 20280–281, collected in Posic (6°26′58.031″ S, 77°14′11.349″ W, WGS84 GPS coordinates) at 1902 m a.s.l., Province Mariscal Cáceres, Departmento San Martín, Peru, on 7 December 2018 by P.J. Venegas. CORBIDI 20300–20305, 20383–20384, collected in Nuevo Chirimoto (6°34′5.716″ S, 77°15′26.49″ W, WGS84 GPS coordinates) at 2019 m a.s.l., Province Mariscal Cáceres, Departmento San Martín, Peru, on 10 December 2018 by L.A. García-Ayachi. CORBIDI 20748, a juvenile, collected in Cataratas de Nueva Esperanza (6°34′7.111″ S, 77°15′5.671″ W, WGS84 GPS coordinates) at 1989 m a.s.l., Province Bagua, Departmento Amazonas, Peru, on 11 August 2019 by P.J. Venegas.
Diagnosis: A large species of Rhinella tentatively assigned to the festae species Group due to having preorbital crest absent, supraorbital crest weak, nuptial pads dark, and tarsal fold absent, characterized by the following combination of characters: (1) SVL 56.1–70.8 mm in males (n = 4), SVL 90.6–91.6 in females (n = 2); (2) snout protruding beyond the margin of lip, slightly pointed in dorsal view, inclined posteroventrally in profile; (3) nostrils protuberant, directed dorsolaterally, anterior half exceeding anterior margin of lower jaw; (4) canthal crest distinct, slightly elevated, supraorbital and postorbital crests weakly defined, covered with tubercles, supratympanic crests indistinguishable, pretympanic crest absent; (5) tympanic membrane and tympanic annulus absent; (6) bony protrusion at angle of jaw absent; (7) neural crest of vertebrae absent; (8) parotoid glands long, prominent, from posterior margin of upper eyelid to posterior level of arm insertion, outside nearly straight, inside bulbous, incorporated into lateral row of tubercles; (9) lateral row of tubercles diagonally from posterior margin of parotoid gland slightly exceeding middle of flanks; tubercles large, rounded to subconical, and widely spaced; (10) skin on dorsum smooth with large scattered round to ovoid subconical tubercles; (11) skin on dorsal surfaces of limbs smooth, tubercular with small scattered round to ovoid subconical tubercles; (12) first finger slightly longer than the second; (13) palmar tubercle large, ovoid, two times size of ovoid thenar tubercle; (14) inner metatarsal tubercle ovoid, barely elevated, two times size of outer round to ovoid subconical metatarsal tubercle; (15) modal webbing on foot: I 1—1+ II 1—3+ III 1—4– IV 4—2+ V in males; (16) subarticular tubercles prominent, round to oval; supernumerary tubercles round, slightly smaller than the former; (17) subgular vocal sac and vocal slits absent, brown nuptial excrescences present on first and second finger in males; (18) in life, dorsum uniform dark purple brown or tan, flanks pale purple brown to tan; venter light grey with brown mottling; iris golden with irregular dark brown mottling.
Comparisons: Morphologically, R. moralesi sp. nov. differs from all members of the festae species Group by having larger males (largest SVL in males of R. moralesi sp. nov. 70.8 mm, R. chavin 52.0 mm [8], R. multiverrucosa 47.7 mm [16], R. lilyrodriguezae 46.4 mm (n = 5), R. chullachaki 44.2 mm [14], R. macrorhina 43.4 mm [10], R. acrolopha 41.9 mm [10], R. yanachaga 41.6 mm [13], R. paraguas 41.7 mm [18], R. rostrata 41 mm [12], R. tenrec 40.2 mm [21], R. ruizi 39.4 mm [19], R. arborescandens 35.3 mm [4], R. festae 33.9 mm [32], R. manu 32.3 mm [5], R. nicefori 32 mm [10], R. tacana 30.6 mm [20], and R. lindae 29.9 mm [15]; males of R. nesiotes, and R. truebae are unknown, but the SVL of females is smaller, 29.9 mm [11] and 65.9 mm [21], respectively vs. 91.6 mm in R. moralesi sp. nov., and by having the outer dorsolateral tarsus surface with a subconical ridge of fused tubercles.
Furthermore, R. moralesi sp. nov. differs from nine members of the R. festae species Group (R. chavin, R. lilyrodriguezae, R. lindae, R. manu, R. multiverrucosa, R. nesiotes, R. tacana, R. truebae, and R. yanachaga) that have an externally visible tympanic annulus and tympanic membrane, whereas it shares with 10 members (R. acrolopha, R. arborescandens, R. chulluchaki, R. festae, R. macrorhina, R. nicefori, R. paraguas, R. rostrata, R. ruizi, and R. tenrec) the absence of an externally visible tympanic annulus and tympanic membrane.
From the latter 10 species, R. moralesi sp. nov. can be distinguished as follows: Rhinella moralesi sp. nov. and R. acrolopha have males that lack vocal slits, but males of R. moralesi sp. nov. are much larger (70.8 mm vs. 41.9 mm) [10]. Furthermore, R. moralesi sp. nov. has prominent elongate parotid glands in contact with the eye (small, ovoid, not in contact with eye). Rhinella moralesi sp. nov. differs from R. arborescandens by having parotoid glands elongate and in contact with the eye (ovoid, not in contact), and males with finger one and two with nuptial pads (distinct keratinous spines on finger one) [4]. Rhinella moralesi sp. nov. differs from R. chullachaki by having parotoid glands elongate and in contact with the eye (ovoid, not in contact), and pretympanic crests absent (well defined) [14]. Rhinella moralesi sp. nov. and R. festae have males that lack vocal slits. However, R. moralesi sp. nov. differs from R. festae by having a short protuberant snout (long) and large dorsal tubercles (minute) [10]. Rhinella moralesi sp. nov. differs from R. macrorhina by having males without vocal slits (present), by having a short protuberant snout (long), a dorsolateral row of widely spaced large, rounded to subconical tubercles (continuous row of small and depressed tubercles), and pretympanic crest absent (present) [10]. Rhinella moralesi sp. nov. differs from R. nicefori by having males without vocal slits (present) and supraorbital crests weakly defined (distinct) [17]. Rhinella moralesi sp. nov. differs from R. paraguas by having parotoid glands in contact with the eye (not in contact) and pretympanic crests absent (well defined) [18]. Rhinella moralesi sp. nov. shares with R. rostrata a row of large dorsolateral tubercles but differs by having long prominent parotoid glands (small, sub-triangular), pretympanic crest absent (present), hands and feet with relatively short and stubby digits (long and slender), and first finger slightly longer than second (first finger about half the size of the second) [12]. Rhinella moralesi sp. nov. differs from R. ruizi by having males with nuptial pads (absent), parotoid glands elongate and in contact with the eye (triangular, not in contact), and long fingers and toes (short) [19]. Rhinella moralesi sp. nov. differs from R. tenrec by lacking a snout with a proboscis (proboscis present), parotoid glands long, prominent, and in contact with the eye (small, triangular, not in contact), and dorsum smooth with large scattered, round to ovoid tubercles (smooth with low tubercles) [21].
Ten species (including the new species) of the Rhinella festae Group are currently recognized in Peru. From the seven species that have an externally visible tympanum, R. moralesi sp. nov. can be distinguished as follows: Rhinella moralesi sp. nov. and R. chavin (Figure 3A,B) share a stout body with relatively short extremities and short digits, large glands on the dorsum and a lateral row of enlarged, elevated glands, males with nuptial pads on the first and second finger and absence of vocal slits [8]. However, R. moralesi sp. nov. has a protuberant, slightly pointed, and posteroventrally inclined snout (truncate in dorsal view, rounded in profile, not protuberant), has the parotoid gland in contact with the eye (narrowly separated), lacks large and elongate glands on the forearm and tibia (present), and lacks keratin-tipped tubercles (present). Rhinella moralesi sp. nov. and R. multiverrucosa (Figure 3A,D) share a stout body with relatively short extremities and short digits, parotoid glands in contact with the eye, large glands on the dorsum, and a lateral row of enlarged, elevated glands, males with nuptial pads on the first and second finger and vocal slits absent [16]. However, R. moralesi sp. nov. has elongated parotoid glands (subtriangular), lacks large glands on the head between parotoid glands (two large glands present), dorsum without keratin-tipped tubercles (present), lacks large glands on forearm and tibia (present), has a protuberant, slightly pointed, and posteroventrally inclined snout (snout truncate in dorsal view, rounded in profile, not protuberant). Rhinella moralesi sp. nov. and R. yanachaga (Figure 3A,C) both have a slightly pointed snout in dorsal view, which is protruding in lateral view, a distinct canthus rostralis, and males have nuptial pads on first and second fingers and a protuberant cloaca with its opening directed ventrally [13]. However, R. moralesi sp. nov. is larger with an SVL of 70.8 mm (41.6 mm), has elongate parotoids in contact with the eye (subtriangular, not in contact with the eye), dorsolateral tubercle row distinct (weakly defined), dorsum in males smooth with large scattered round to ovoid subconical tubercles (numerous small keratin-tipped tubercles), and males lack vocal slits (present). Rhinella moralesi sp. nov. is easily distinguished from R. lilyrodriguezae by having relatively short and stubby extremities (long and slender in R. lilyrodriguezae), having a short snout (long), lacking pretympanic crests (present), and dorsum with large glands (small glands) [7]. Rhinella moralesi sp. nov. and R. manu have males without vocal slits and a slightly pointed and protruding snout [5]. However, R. moralesi sp. nov. has the parotoid gland contacting the eye (not in contact), nuptial pads on fingers one and two (distinct keratinous spines on finger one), and dorsum with large glands (with numerous small glands). Rhinella moralesi sp. nov. can be distinguished from R. tacana by lacking a visible tympanum (present), males without vocal slits (present), dorsum with large glands (absent), and a maximum SVL in males of 70.8 mm (30.7 mm) [20]. Rhinella moralesi sp. nov. and R. nesiotes have the snout pointed in dorsal view and projecting. However, R. moralesi sp. nov. has a lateral row of tubercles (absent), cranial crests present (absent), and the parotoid gland is elongated and contacting the eye (ovoid, not contacting) [11].
Description of holotype: Adult male; body robust; SVL 67.6 mm; head about as wide as long; snout slightly pointed in dorsal view, protruding beyond the margin of lip, rounded above and inclined posteroventrally in profile (Figure 5B), with a broad fleshy vertical keel; canthus rostralis nearly straight in dorsal view, angular in profile; canthal crest distinct, slightly elevated, supraorbital and postorbital crests weakly defined, covered with tubercles, supratympanic crests indistinguishable, pretympanic crest absent; dorsum of head flat, skin co-ossified with underlying cranial bones; interorbital distance slightly larger than eyelid width; horizontal eye diameter slightly larger than distance between nostril and anterior corner of eye; internarial area concave; nostrils protuberant, directed laterally, anterior half exceeding anterior margin of lower jaw; loreal region strongly concave with tiny subconical granules; lips rounded bearing tiny subconical granules; small V-shaped notch at symphysis of upper jaw; tympanic membrane and tympanic annulus absent; tympanic area with tubercular folds and tiny subconical granules; corner of upper jaws with two prominent conical tubercles on each side (Figure 5B); bony protrusion at angle of jaw absent; neural crest of vertebrae absent; skin on dorsum and flanks smooth with large scattered round to ovoid subconical tubercles lacking keratinized tips; parotoid glands long, prominent, from posterior margin of upper eyelid to posterior level of arm insertion, outside nearly straight, inside bulbous; posterior end of parotoid gland incorporated into lateral row of widely spaced rounded to subconical tubercles extending diagonally and slightly exceeding middle of flanks (Figure 3A); skin on dorsal surfaces of limbs smooth, tubercular with scattered round to ovoid subconical tubercles slightly smaller than those on dorsum; cranial crests and parotoid glands smooth; upper eye lids with a prominent conical outer ridge and round to ovoid subconical tubercles; skin on throat, chest, belly, and ventral surfaces of forelimbs and hind limbs coarsely areolate to warty (Figure 4B); forelimb relatively long; fingers with lateral fringes; tips of digits rounded, terminating in indistinct discs; relative length of fingers I > II < IV< III; palmar tubercle large, ovoid, two times size of ovoid thenar tubercle; subarticular tubercles most prominent at the base of fingers, round to oval, distal subarticular tubercles on Finger II on left hand and Finger IV on right hand bifid; supernumerary tubercles numerous, about half size of subarticular tubercles; basal webbing between fingers; brown nuptial excrescences present on dorsal surfaces of fingers I and II (Figure 5C,D); foot longer than tibia; relative length of toes I < II < III = V < IV; inner tarsal fold absent; outer dorsolateral tarsus surface with a subconical ridge of fused tubercles (Figure 4A); inner metatarsal tubercle ovoid, barely elevated, two times size of outer round to ovoid subconical metatarsal tubercle; subarticular tubercles most prominent at the base of toes, round to oval, distal subarticular tubercles often irregularly shaped or bifid; supernumerary tubercles round, slightly smaller than subarticular tubercles; toes with moderate webbing, I 1–1+ II 1––3+ III 1–4– IV 4––2+ V; lateral fringes broad; tips of digits rounded, terminating in indistinct discs (Figure 5E); tongue elongate, two thirds attached to mouth floor; choanae small, oval; dentigerous processes of vomers and maxillary teeth absent; vocal slits absent; subgular vocal sac absent.
Measurements (in mm) of the holotype: SVL 67.6; TL 26.8; FL 28.2; HL 23.7; HW 23.9; ED 5.6; IOD 6.9; EW 6.7; IND 4.6; E-N 5.2; PL 17.7; PW 6.4.
Coloration of holotype in life (Figure 3A): Dorsum dark purple-brown, flanks pale purple-brown to tan; iris golden with irregular dark brown mottling.
Coloration of holotype in alcohol (Figure 4A,B, and Figure 5): Dorsal surfaces pale hazel brown with tubercles and parotid glands grayish brown. Middorsal tan hairline from snout to cloaca. Nuptial pads hazel brown and distinct from the grayish tan coloration of hands (Figure 4D). Lateral surfaces of head (Figure 4B) tan with pale hazel brown mottling. Flanks colored as dorsum. Throat, belly, and ventral surfaces of legs grayish tan with pale brown mottling. Ventral surfaces of hands and feet pale gray (Figure 5C–E).
Variation: For measurements of adult paratypes see Appendix B (Table A1) and for ranges and proportions of adult type specimens see Appendix C (Table A2).
All paratypes (Figure 3C,D, Figure 6 and Figure 7) are similar to the holotype regarding morphology and proportions. Females are larger than males. All adult males have brown nuptial pads and lack a subgular vocal sac. Adults have a ridge of fused tubercles on the outer dorsolateral surface of the tarsus (Figure 6A and Figure 7A). The smallest juvenile (CROBIDI 20281) has an SVL of 17.7 mm, the largest 29.3 mm (CORBIDI 20305). The coloration for a male (MUSM 15958, Figure 3C,D) in life was noted by AC as dark tan dorsally, without patterns, and light grey with brown mottling ventrally. In alcohol, the paratype is colored like the holotype except for a slightly broader middorsal hairline (Figure 3C). Dorsal coloration in adults in life is dark brown (CORBIDI 20372) or pale brown (CORBIDI 713) and the venter is pale grayish cream with brown (CORBIDI 20372) or gray (CORBIDI 713) marmoration. Hand and toe surfaces are dark gray (CORBIDI 713, 20371) or pale gray (CORBIDI 20372) with toe and fingertips ranging from dark gray (CORBIDI 713) to brownish orange (CORBIDI 20371). One juvenile is blackish brown (CORBIDI 18862) dorsally and laterally and has the parotoid gland with pale olive flecks and the lateral line of tubercles is pale olive (Figure 6G), the venter is slightly paler with blackish-brown blotches. One juvenile (CORBIDI 18862) is dorsally, laterally, and ventrally dark brown and orange-brown marmorated. The parotoid gland, lateral line of tubercles, and groin are tan.
Etymology: We dedicate the new species to our late colleague and friend Professor Victor Morales in recognition of his contributions to Neotropical herpetology.
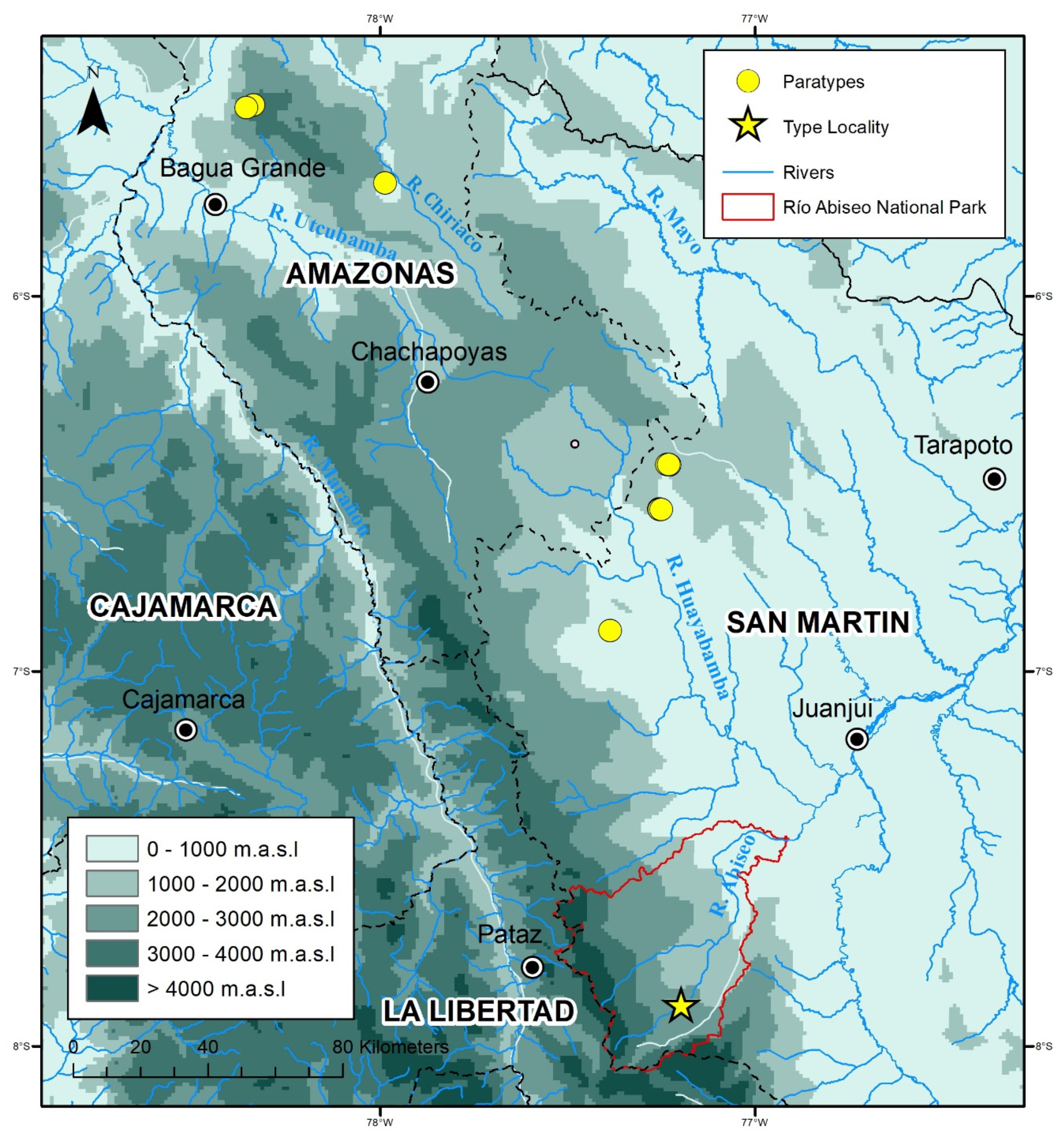
Distribution, ecology, and threat status: Rhinella moralesi sp. nov. is known from seven montane forest localities between 1788 and 2305 m a.s.l. in Departamentos Amazonas and San Martín (Figure 1 and Figure 2). The distance between the northernmost and southernmost distribution records for R. moralesi sp. nov. is about 294 km (airline). At Río Abiseo National Park, both specimens were found in proximity: the holotype was found in a terrestrial Sherman trap, and the male paratype (MUSM 15958) was found by botanists under unknown circumstances. The type locality was characterized by very humid montane forest, with some clearings and disturbed areas. There were signs of past and current burning along most of the Río Abiseo Valley sites that AC and LOR visited, including Jucusbamba. Eight 10 × 10 m2 leaf litter plots searched at elevations from 1800 to 2000 m a.s.l. around Jucusbamba and the nearby locality of Granadilla, and two nocturnal transects in Jucusbamba (1730–1800 m a.s.l.) did not reveal any amphibians. At Granadilla (1990–2070 m a.s.l.), amphibians (possibly sympatric with R. moralesi sp. nov.) observed during four nocturnal transects included Gastrotheca sp. gr. testudinea, Pristimantis cf. corrugatus, and Pristimantis sp.
At other sites in northern Peru, one adult female specimen (CORBIDI 713) was found inactive under a log during the day at 1100 h. Two specimens (male CORBIDI 20268, subadult female CORBIDI 20837) were found walking on the leaf litter by night between 2000 and 2200 h. One couple (male CORBIDI 20371, female CORBIDI 20372, Figure 7A) were found in amplexus walking on the leaf litter at 1000 h on 10 December 2018. All juveniles were found at night perched on leaves between 10 or 30 cm above ground or on the leaf litter. Specimens collected in Laguna Negra, Cataratas de Nueva Esperanza, and Uriarte (CORBIDI 713, 20748, 20837) were collected in primary montane forest, while specimens from Yambrasbamba, Posic, and Nuevo Chirimoto were collected in patches of montane forest near cropland of corn (Zea mays) and pastures for cattle ranching. Subadult and adult specimens of R. moralesi sp. nov. did not show saltatorial locomotion when captured or when handled for photography, while some juveniles showed weak jumps attempting to escape.
In Laguna Negra, Posic, and Nuevo Chirimoto, R. moralesi sp. nov. was collected in sympatry with Gastrotheca testudinea, Hemiphractus sp., Pristimantis nephophilus, and Rhinella margaritifera complex; at Yambrasbamba in sympatry with P. galdi, P. nephophilus, P. schultei, Hyloscirtus phyllognathus, and Nymphargus posadae; at Cataratas de Nueva Esperanza and Uriarte in sympatry with Pristimantis serendipitus, P. katoptroides, P. schultei, P. sp., Lynchius sp., and Noblella sp.
We detected infection by the pathogenic chytrid fungus (Bd) in the only sampled individual (CORBIDI 20748), captured at 1695 m a.s.l. in Nueva Esperanza (Bagua Province, Amazonas Department) on 11 August 2019. The level of infection ZE = 14 zoospores is low and not typically associated with diseased individuals. The presence of Bd in northern Peru has previously been reported, and the oldest confirmed record for the country is from northern Peru, during the 1999 mass die-off of Atelopus patazensis in Pataz, Department of La Libertad [33]. The distribution records of R. moralesi sp. nov. cover a polygon area of 10661.84 km2 (EOO) and the AAO is 32 km2 revealing an IUCN conservation status of either “Vulnerable” or “Endangered”. Based on habitat destruction outside of protected areas resulting from agricultural activities and a Bd record, we consider the threat status for R. moralesi sp. nov. as “Vulnerable” according to the IUCN red list criteria and categories [34].
4. Discussion
Despite the absence of molecular data, the distinct morphological characters facilitated the description of Rhinella moralesi sp. nov. The lack of a visible tympanic annulus and membrane has been noted for various species of Rhinella from different species groups, suggesting an evolutionarily independent loss of these characters. The complex history of the loss of the ear in anurans has been discussed by Pereyra et al. [35] and von May et al. [36]. Pereyra et al. [3] recognized that the Rhinella margaritifera species Group and the R. veraguensis species Group are sister clades of the R. festae species Group. We feel confident with the assignment of R. moralesi sp. nov. to the festae species Group. The nine members of the veraguensis Group [3] have males with light-colored nuptial pads whereas the nuptial pads in members of the festae Group are dark-colored. Furthermore, R. moralesi sp. nov. differs from most members of the veraguensis Group (except R. fissipes, R. justinanoi, R. quechua, and R. veraguensis) by lacking an external visible tympanic annulus and tympanic membrane. However, R. moralesi sp. nov. differs from R. fissipes by having prominent, long parotoid glands (moderately prominent, narrow, [37]) and by having the first finger shorter (equal in length, [37]) than the second, from R. justinanoi by having the first finger shorter than the second (longer, [38], and by having the venter coarsely areolate to warty (venter with spinous, conical tubercles, [38]), and from R. veraguensis by having prominent, long parotoid glands (ovoid, short). The R. margaritifera species Group has 17 species [3]. Rhinella moralesi sp. nov. differs from all by (except R. iserni) by lacking an external visible tympanic annulus and tympanic membrane. Rhinella moralesi sp. nov. differs from R. iserni by having females up to 91.6 mm SVL (65.5 mm, [6]), and by having the first finger shorter (longer, [6]) than the second.
With the description of R. moralesi sp. nov., the festae Group currently has 20 species in Panama, Andean Columbia, Ecuador, Peru, and Bolivia between 200 and 3100 m a.s.l. (Figure 1). The lowest elevation is known for R. festae (200‒1700 m a.s.l.), the highest for R. ruizi (3100 m a.s.l. [19]. Interestingly, the majority of the species of the R. festae Group have been observed above ground in the vegetation, and referred to as semiarboreal or arboreal, often along with lack of saltatory locomotion which is replaced by walking/climbing (e.g., R. chavin, R. yanachaga, see Figure 3). Only four species (R. acrolopha, R. lindae, R. macrorhina, and R. rostrata) are considered terrestrial [10,12,15], whereas the remaining species are described as either arboreal or semiarboreal, except for three species (R. multiverrucosa, R. tenrec, and R. truebae) whose natural history is unknown. We consider R. moralesi sp. nov. semiarboreal, as specimens were found on the ground and 30 cm above the ground in the vegetation.
For the majority of the members of the festae species Group, the reproduction mode remains a mystery because eggs or tadpoles have not been found. However, unpigmented, vitelline-rich eggs have often been noticed during dissections of preserved specimens, except for two species whose females are unknown (e.g., R. chullachaki, R. moralesi sp. nov., and R. rostrata). Lehr et al. [8] noticed vitelline-rich eggs (266‒286, n = 2) in oviducts of R. chavin and suspected direct development; the eggs had a diameter of 1.8 and 3.1 mm (n = 10), respectively. This is supported by the lack of running or temporary waters where R. chavin was found both at the type locality and in the Cordillera de Carpish. Few, large, vitelline-rich eggs have also been noticed in oviducts of R. nesiotes [11], R. manu, R. tacana, and R. yanachaga [13] suggesting they also may reproduce through direct development. Rhinella manu is often found far away from water, including on mountain ridges with no nearby streams. One gravid female of R. lilyrodriguezae (MUSM 32204) was observed perching on palm leaves. Its dissection revealed a total of 185 vitelline-rich eggs with an average diameter of 2.8 mm. Two females of R. cf. manu (CORBIDI 16349, 16360) had 20 and 40 vitelline-rich eggs in their oviducts, on average 1.3 mm and 2.4 mm in diameter, respectively (Catenazzi, unpubl. data).
The large distributional gabs between species of the R. festae Group, most of which are only known from their type localities or close surroundings (Figure 1), suggests that many new species of Rhinella await discovery in the Andean montane forests.
Author Contributions
Conceptualization, E.L.; Funding acquisition, E.L., P.J.V.; Investigation, E.L., J.C.C., P.J.V., L.A.G.-A. and A.C.; Methodology, E.L., J.C.C., L.O.R., P.J.V., L.A.G.-A. and A.C.; Project administration, E.L.; Writing—original draft, E.L. and A.C.; Writing—review & editing, E.L., J.C.C., L.O.R., P.J.V., L.A.G.-A. and A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Illinois Wesleyan University with an ASD grant to EL and with funds from Critical Ecosystem Partnership Fund, with the projects numbers CEPF-108792 and CEPF–109938, and Fondo de Promoción de las Áreas Naturales Protegidas del Perú (PROFONANPE) to PJV.
Institutional Review Board Statement
Ethical review and approval (protocol 19-011) were granted for this study by the IACUC committee of Illinois Wesleyan University.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Specimens were kindly loaned by C. Aguilar (MUSM). EL thanks IWU for an ASD grant to support this research. PJV’s fieldwork was supported by the Critical Ecosystem Partnership Fund, Global Genome Initiative (GGBN–GGI), and NGO’s Ucumari and Rainforest Partnership. PJV’s fieldwork would not have been possible without the logistic support of R. Wagter, F. Rubio, O. Sanchez, M. Leo, and G. Seitz. PJV is especially grateful to his field companions J. Briones, J. Ormeño, A. Marchelie, D. Vasquez, J. Gamboa, and J. C. Chávez. Specimens housed in CORBIDI were collected with the following permits: 110-2007-INRENA-IFFS-DCB; RDG N°299-2017-SERFOR-DGGSPFFS; RDG N°295-2017-SERFOR-DGGSPFFS; and RDG N°067-2019-SERFOR-DGGSPFFS. LOR thanks APECO and the Bio-Andes project; special thanks go to A. Millington. Specimens hold at the MUSM were collected under permit 89-98-INRENA-DGANPFS-DANP. We thank two anonymous reviewers for their helpful feedback that improved our manuscript, and M. Lundberg for his photograph of R. multiverrucosa.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The following specimens were examined: Rhinella chavin (Lehr, Köhler, Aguilar, and Ponce, 2001): Peru: Huánuco: Chaglla, Palmapampa, 20 km southeast of Chaglla, 3010 m a.s.l.: MUSM 20028 (holotype); Rhinella lilyrodriguezae Cusi, Moravec, Lehr, and Gvoždík, 2017: Peru: San Martín: collected ca. 20 km from Park Rangers Center N° 53 “Shapaja” of the Cordillera Azul National Park, 1260 m: MUSM 32204 (holotype); Puesto de Control 16 (Shambirillo), Parque Nacional Cordillera Azul, 1122 m a.s.l.: CORBIDI 8838–42, CORBIDI 9188–93, CORBIDI 9928, CORBIDI 9948–50; Rhinella cf. manu Chaparro, Parmuk, and Gluesenkamp, 2007: Peru: Cusco: Paucartambo: Kosñipata Valley, 1830–2150 m a.s.l.: MUSM 21129, 26282, 27929, 27931–33; CORBIDI 16234–36, 16239, 16349, 16359–60, 19191; Rhinella multiverrucosa (Lehr, Pramuk, and Lundberg, 2005): Peru: Pasco: NW Auquimarca, 2900 m a.s.l.: MUSM 17820 (holotype); Rhinella tacana (Padial, Reichle, McDiarmid, and De la Riva, 2006): Peru: Cusco: Paucartambo: Chontachaca, Kosñipata Valley, 915 m a.s.l.: CORBIDI 19184; Rhinella yanachaga Lehr, Pramuk, Hedges, and Córdova 2007: Peru: Pasco: Oxapampa: Quebrada Yanachaga, 2900 m a.s.l.: FMNH 282819, MUSM 31100.
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Measurements (in mm) of adult male and female paratypes of Rhinella moralesi sp. nov.
| Characters | CORBIDI | CORBIDI | CORBIDI | MUSM | CORBIDI |
|---|---|---|---|---|---|
| 20372 | 713 | 20371 | 15958 | 20268 | |
| Sex | F | F | M | M | M |
| SVL | 91.6 | 90.6 | 70.8 | 57.6 | 56.1 |
| TL | 39.2 | 37.5 | 30.7 | 22.7 | 21.9 |
| FL | 39.8 | 37.8 | 32.3 | 23.9 | 22.2 |
| HL | 29.1 | 29.6 | 23 | 18.2 | 16.9 |
| HW | 35.8 | 33.6 | 26.4 | 19.8 | 22 |
| ED | 7.8 | 6.7 | 6.1 | 4.9 | 6 |
| IOD | 14.2 | 12.4 | 9.6 | 6.3 | 8.3 |
| EW | 6.6 | 7.4 | 5.3 | 5.8 | 5.2 |
| IND | 4.7 | 5 | 3.8 | 3.4 | 3.6 |
| E–N | 8.1 | 7.7 | 6 | 4.9 | 5 |
| PL | 27.3 | 25.9 | 18 | 13.6 | 15.9 |
| PW | 12.4 | 9.9 | 7.4 | 14.1 | 5.8 |
Appendix C
Table A2.
Ranges (in mm) and proportions of adult male and female types of Rhinella moralesi sp. nov.; ranges followed by means and one standard deviation in parentheses.
Table A2.
Ranges (in mm) and proportions of adult male and female types of Rhinella moralesi sp. nov.; ranges followed by means and one standard deviation in parentheses.
| Characters | Males (n = 4) | Females (n = 2) |
|---|---|---|
| SVL | 56.1–70.8 (63.0 ± 7.3) | 90.6–91.6 (91.1 ± 0.7) |
| TL | 21.9–30.7 (25.5 ± 4.1) | 37.5–39.2 (38.4 ± 1.2) |
| FL | 22.2–32.3 (26.7 ± 4.5) | 37.8–39.8 (38.8 ± 1.4) |
| HL | 16.9–23.7 (20.5 ± 3.4) | 29.1–29.6 (29.4 ± 0.4) |
| HW | 19.8–26.4 (23.0 ± 2.8) | 33.6–35.8 (34.7 ± 1.6) |
| ED | 4.9–6.1 (5.7 ± 0.5) | 6.7–7.8 (7.3 ± 0.8) |
| IOD | 6.3–9.6 (7.8 ± 1.5) | 12.4–14.2 (13.3 ± 1.3) |
| EW | 5.2–6.7 (5.8 ± 0.7) | 6.6–7.4 (7.0 ± 0.6) |
| IND | 3.4–4.6 (3.9 ± 0.5) | 4.7–5.0 (4.9 ± 0.2) |
| E–N | 4.6–5.6 (5.1 ± 0.4) | 7.7–8.1 (7.9 ± 0.3) |
| PL | 13.6–18.0 (16.3 ± 2.0) | 25.9–27.3 (26.6 ± 1.0) |
| PW | 5.8–14.1 (8.4 ± 3.8) | 9.9–12.4 (11.2 ± 1.8) |
| TL/SVL | 0.39–0.43 | 0.41–0.43 |
| FL/SVL | 0.40–0.46 | 0.42–0.43 |
| HL/SVL | 0.30–0.35 | 0.32–0.33 |
| HW/SVL | 0.34–0.39 | 0.37–0.39 |
| HW/HL | 1.01–1.15 | 1.14–1.23 |
| E–N/ED | 0.77–1.00 | 1.04–1.15 |
| EW/IOD | 0.55–0.97 | 0.46–0.60 |
| PL/PW | 0.96–2.77 | 2.20–2.62 |
References
- Fitzinger, L.J. Neue Classification der Reptilien nach ihren Natürlichen Verwandtschaften nebst einer Verwandtschafts-Tafel und einem Verzeichnisse der Reptilien-Sammlung des K. K. Zoologisch Museum’s zu Wien; J.G. Heubner: Vienna, Austria, 1826; pp. 1–66. [Google Scholar]
- Frost, D. Amphibian Species of the World: An Online Reference. Version 6.1. Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 18 June 2021).
- Pereyra, M.O.; Blotto, B.L.; Baldo, D.; Chaparro, J.C.; Ron, S.R.; Elias-Costa, A.J.; Iglesias, P.P.; Venegas, P.J.; Thomé, M.T.C.; Ospina-Sarria, J.J.; et al. Evolution in the Genus Rhinella: A total evidence phylogenetic analysis of neotropical true toads (Anura: Bufonidae). Bull. Am. Mus. Nat. Hist. 2021, 447, 1–156. [Google Scholar] [CrossRef]
- Duellman, W.E.; Schulte, R. Description of a new species of Bufo from northern Peru with comments on phenetic groups of South American toads (Anura: Bufonidae). Copeia 1992, 1992, 162–172. [Google Scholar] [CrossRef]
- Chaparro, J.C.; Pramuk, J.B.; Gluesenkamp, A.G. A new species of arboreal Rhinella (Anura: Bufonidae) from cloud forest of southeastern Peru. Herpetologica 2007, 63, 203–212. [Google Scholar] [CrossRef]
- Moravec, J.; Lehr, E.; Cusi, J.C.; Córdova, J.H.; Gvoždík, V. A new species of the Rhinella margaritifera species Group (Anura, Bufonidae) from the montane forest of the Selva Central, Peru. ZooKeys 2014, 371, 35–56. [Google Scholar] [CrossRef] [Green Version]
- Cusi, J.C.; Moravec, J.; Lehr, E.; Gvoždík, V. A new species of semiarboreal toad of the Rhinella festae Group (Anura, Bufonidae) from the Cordillera Azul National Park, Peru. ZooKeys 2017, 673, 21–47. [Google Scholar] [CrossRef] [Green Version]
- Lehr, E.; Köhler, G.; Aguilar, C.; Ponce, E. New species of Bufo (Anura: Bufonidae) from central Peru. Copeia 2001, 2001, 216–223. [Google Scholar] [CrossRef]
- Peracca, M.G. Viaggio del Dr. Enrico Festa nell’ Ecuador e Regioni Vicine. Rettili ed Anfibi. In Bollettino dei Musei di Zoologia e Anatomia Comparata della R; Universita di Torino: Torino TO, Italy, 1904; Volume 19, pp. 1–41. [Google Scholar]
- Trueb, L. Phylogenetic Relationships of Certain Neotropical Toads with the Description of a New Genus (Anura: Bufonidae); Contributions in Science; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1971; Volume 216, pp. 1–40. [Google Scholar]
- Duellman, W.E.; Toft, C.A. Anurans from the Serranía de Sira, Amazonian Perú: taxonomy and biogeography. Herpetologica 1979, 35, 60–70. [Google Scholar]
- Noble, G.K. Two new batrachians from Colombia. Bull. Am. Mus. Nat. Hist. 1920, 42, 441–446. [Google Scholar]
- Lehr, E.; Pramuk, J.B.; Hedges, S.B.; Córdova, J.H. A new species of arboreal Rhinella (Anura: Bufonidae) from Yanachaga-Chemillén National Park in central Peru. Zootaxa 2007, 1662, 1–14. [Google Scholar]
- Castillo-Urbina, E.; Glaw, F.; Aguilar-Puntriano, C.; Vences, M.; Köhler, J. Genetic and morphological evidence reveal another new toad of the Rhinella festae species group (Anura: Bufonidae) from the Cordillera Azul in central Peru. Salamandra 2021, 57, 181–195. [Google Scholar]
- Rivero, J.A.; Castaño, C.J. A new and peculiar species of Rhamphophryne (Amphibia: Bufonidae) from Antioquia, Colombia. J. Herpetol. 1990, 24, 1–5. [Google Scholar] [CrossRef]
- Lehr, E.; Pramuk, J.B.; Lundberg, M. A new species of Bufo (Anura: Bufonidae) from Andean Peru. Herpetologica 2005, 61, 308–318. [Google Scholar] [CrossRef]
- Cochran, D.M.; Goin, J.C. Frogs of Colombia. Bull. U.S. Natl. Mus. 1970, 288, 1–655. [Google Scholar] [CrossRef]
- Grant, T.; Bolívar-G, W. A new species of semiarboreal toad with a salamander-like ear (Anura: Bufonidae: Rhinella). Herpetologica 2014, 70, 198–210. [Google Scholar] [CrossRef]
- Grant, T. Una nueva especie de Rhamphophryne (Anura: Bufonidae) de la Cordillera Central de Colombia. Rev. Acad. Colomb. Cienc. Exact. Fis. Nat 1999, 23, 287–292. [Google Scholar]
- Padial, J.M.; Reichle, S.; McDiarmid, R.W.; De la Riva, I. A new species of arboreal toad (Anura: Bufonidae: Chaunus) from Madidi National Park, Bolivia. Zootaxa 2006, 1278, 57–58. [Google Scholar] [CrossRef]
- Lynch, J.D.; Renjifo, J.M. Two new toads (Bufonidae: Rhamphophryne) from the Northern Andes of Colombia. J. Herpetol. 1990, 24, 364–371. [Google Scholar] [CrossRef]
- Rodríguez, L.O.; Catenazzi, A. Four new species of terrestrial-breeding frogs of the genus Phrynopus (Anura: Terrarana: Craugastoridae) from Río Abiseo National Park, Peru. Zootaxa 2017, 4273, 381–406. [Google Scholar] [CrossRef]
- Lehr, E.; Gregory, C.; Catenazzi, A. A new species of Pristimantis (Amphibia: Anura: Strabomantidae) from the Río Abiseo National Park, Peru. Zootaxa 2013, 3731, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Lehr, E.; Catenazzi, A. A new species of marsupial frog (Anura: Hemiphractidae: Gastrotheca) from the Río Abiseo National Park in Peru. Herpetologica 2011, 67, 449–459. [Google Scholar] [CrossRef]
- Myers, C.W.; Duellman, W.E. A new species of Hyla from Cerro Colorado, and other tree frog records and geographical notes from Western Panama. Am. Mus. Novit. 1982, 2752, 1–32. [Google Scholar]
- Duellman, W.E. The Hylid Frogs of Middle America; Monograph of the Museum of Natural History; The University of Kansas: Lawrence, KS, USA, 1970; Volume 1, pp. 1–753. [Google Scholar]
- Duellman, W.E.; Lehr, E. 2009. Terrestrial-Breeding Frogs (Strabomantidae) in Peru; Natur und Tier-Verlag, Naturwissenschaft: Münster, Germany, 2009; pp. 1–382. [Google Scholar]
- Duellman, W.E.; Mendelson, J.R. Amphibians and reptiles from northern Departamento Loreto, Peru: taxonomy and biogeography; The University of Kansas Science Bulletin: Lawrence, KS, USA, 1995; Volume 55, pp. 329–376. [Google Scholar]
- Lötters, S.; Köhler, J. A new toad of the Bufo typhonius complex from humid montane forests of Bolivia (Amphibia, Anura, Bufonidae). Spixiana 2000, 23, 293–303. [Google Scholar]
- Hyatt, A.D.; Boyle, D.G.; Olsen, V.; Boyle, D.B.; Berger, L.; Obendorf, D.; Dalton, A.; Kriger, K.; Hero, M.; Hines, H.; et al. Diagnostic assays and sampling protocols for the detection of Batrachochytrium dendrobatidis. Dis. Aquat. Organ. 2007, 73, 175–192. [Google Scholar] [CrossRef]
- Boyle, D.G.; Boyle, D.B.; Olsen, V.; Morgan, J.A.T.; Hyatt, A.D. Rapid quantitative detection of chytridiomycosis (Batrachochytrium Dendrobatidis) in amphibian samples using real-time taqman PCR assay. Dis. Aquat. Organ. 2004, 60, 141–148. [Google Scholar] [CrossRef]
- Duellman, W.E.; Lynch, J.D. Anuran amphibians from the Cordillera de Cutucu, Ecuador. Proc. Acad. Nat. Sci. USA Philadelphia 1988, 140, 125–142. [Google Scholar]
- Venegas, P.; Catenazzi, A.; Siu Ting, K.; Carrillo, J. Two new species of harlequin frogs (Anura: Bufonidae: Atelopus) from the Andes of northern Peru. Salamandra 2008, 44, 163–176. [Google Scholar]
- IUCN Standards and Petitions Subcommittee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 10.1. Prepared by the Standards and Petitions Subcommittee. 2013. Available online: http://www.iucnredlist.org/documents/RedListGuidelines.pdf. (accessed on 7 July 2021).
- Pereyra, M.O.; Womack, M.C.; Barrionuevo, J.S.; Blotto, B.L.; Baldo, D.; Targino, M.; Ospina-Sarria, J.J.; Guayasamin, J.M.; Coloma, L.A.; Hoke, K.L.; et al. The complex evolutionary history of the tympanic middle ear in frogs and toads (Anura). Sci. Rep. 2016, 6, 34130. [Google Scholar] [CrossRef] [Green Version]
- von May, R.; Lehr, E.; Rabosky, D. Evolutionary radiation of earless frogs in the Andes: molecular phylogenetics and habitat shifts in high-elevation terrestrial beeding frogs. PeerJ 2018, 6, e4313. [Google Scholar] [CrossRef] [Green Version]
- Boulenger, G.A. Descriptions of new batrachians in the British Museum. Ann. Mag. Nat. Hist. 1903, 12, 552–557. [Google Scholar] [CrossRef]
- Harvey, M.B.; Smith, E.N. A new species of Bufo (Anura: Bufonidae) from cloud forests in Bolivia. Herpetologica 1994, 50, 32–38. [Google Scholar]
Figure 1.
Map indicating type localities (except for R. truebae) of the members of the Rhinella festae species Group. The yellow star indicates the new species. 1. R. acrolopha, 2. R. lindae, 3. R. tenrec, 4. R. nicefori, 5. R. rostrata, 6. R. ruizi, 7. R. macrorhina, 8. R. paraguas, 9. R. festae, 10. R. arborescandens, 11. R. moralesi sp. nov., 12. R. lilyrodriguezae, 13. R. chullachaki, 14. R. chavin, 15. R. nesiotes, 16. R. multiverrucosa, 17. R. yanachaga, 18. R. manu, 19. R. tacana. GIS Map by J.C. Cusi.
Figure 1.
Map indicating type localities (except for R. truebae) of the members of the Rhinella festae species Group. The yellow star indicates the new species. 1. R. acrolopha, 2. R. lindae, 3. R. tenrec, 4. R. nicefori, 5. R. rostrata, 6. R. ruizi, 7. R. macrorhina, 8. R. paraguas, 9. R. festae, 10. R. arborescandens, 11. R. moralesi sp. nov., 12. R. lilyrodriguezae, 13. R. chullachaki, 14. R. chavin, 15. R. nesiotes, 16. R. multiverrucosa, 17. R. yanachaga, 18. R. manu, 19. R. tacana. GIS Map by J.C. Cusi.

Figure 2.
Map with the distributional records of Rhinella moralesi sp. nov. in Departamentos Amazonas and San Martín. GIS Map by J.C. Cusi.
Figure 2.
Map with the distributional records of Rhinella moralesi sp. nov. in Departamentos Amazonas and San Martín. GIS Map by J.C. Cusi.

Figure 3.
Life specimens in dorsolateral views of (A) Rhinella moralesi sp. nov. (holotype, MUSM 15959, SVL 67.6 mm, male), (B) R. chavin (MUSM 20027, SVL 64.9, female) (C) R. yanachaga (MUSM 31100, juvenile), and (D) R. multiverrucosa (MTD 44749, SVL 68.9 mm, female). Photos by A. Catenazzi, (A) E. Lehr (B,C), and M. Lundberg (D).
Figure 3.
Life specimens in dorsolateral views of (A) Rhinella moralesi sp. nov. (holotype, MUSM 15959, SVL 67.6 mm, male), (B) R. chavin (MUSM 20027, SVL 64.9, female) (C) R. yanachaga (MUSM 31100, juvenile), and (D) R. multiverrucosa (MTD 44749, SVL 68.9 mm, female). Photos by A. Catenazzi, (A) E. Lehr (B,C), and M. Lundberg (D).

Figure 4.
Holotype of Rhinella moralesi sp. nov. (MUSM 15959) in (A) dorsal, (B) ventral, and paratype of R. moralesi sp. nov. (MUSM 15958, SVL 57.6 mm, male) in (C) dorsal and (D) ventral views. Red arrows indicate ridge of fused tubercles on outer dorsolateral surface of tarsus. Photos by E. Lehr.
Figure 4.
Holotype of Rhinella moralesi sp. nov. (MUSM 15959) in (A) dorsal, (B) ventral, and paratype of R. moralesi sp. nov. (MUSM 15958, SVL 57.6 mm, male) in (C) dorsal and (D) ventral views. Red arrows indicate ridge of fused tubercles on outer dorsolateral surface of tarsus. Photos by E. Lehr.

Figure 5.
Rhinella moralesi sp. nov. (MUSM 15959, holotype). Head in (A) dorsal and (B) lateral views, hand in (C) ventral, (D) dorsal, and foot in (E) ventral views. Photos by E. Lehr.
Figure 5.
Rhinella moralesi sp. nov. (MUSM 15959, holotype). Head in (A) dorsal and (B) lateral views, hand in (C) ventral, (D) dorsal, and foot in (E) ventral views. Photos by E. Lehr.

Figure 6.
Adult females and juveniles of Rhinella moralesi sp. nov. Female (CORBIDI 20372, SVL 91.6 mm) in dorsal (A) and ventral (B) views, female (CORBIDI 713, SVL 90.6 mm) in dorsal (E) and ventral (F) views. Juvenile (CORBIDI 20301, SVL 26.9 mm) in lateral (C) and ventral (D) views, and juvenile (CORBIDI 18862, SVL 41.3 mm) in lateral (G) and ventral (H) views. Photos by P.J. Venegas (E–H) and L.A. García-Ayachi (A–D).
Figure 6.
Adult females and juveniles of Rhinella moralesi sp. nov. Female (CORBIDI 20372, SVL 91.6 mm) in dorsal (A) and ventral (B) views, female (CORBIDI 713, SVL 90.6 mm) in dorsal (E) and ventral (F) views. Juvenile (CORBIDI 20301, SVL 26.9 mm) in lateral (C) and ventral (D) views, and juvenile (CORBIDI 18862, SVL 41.3 mm) in lateral (G) and ventral (H) views. Photos by P.J. Venegas (E–H) and L.A. García-Ayachi (A–D).

Figure 7.
Male (CORBIDI 20371, SVL 70.8 mm) and female (CORBIDI 20372, SVL 91.6 mm) Rhinella moralesi sp. nov. in amplexus (A), male (CORBIDI 20371) in ventral view (B). Photos by L. A. García-Ayachi.
Figure 7.
Male (CORBIDI 20371, SVL 70.8 mm) and female (CORBIDI 20372, SVL 91.6 mm) Rhinella moralesi sp. nov. in amplexus (A), male (CORBIDI 20371) in ventral view (B). Photos by L. A. García-Ayachi.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lehr, E.; Cusi, J.C.; Rodriguez, L.O.; Venegas, P.J.; García-Ayachi, L.A.; Catenazzi, A. A New Species of Toad (Anura: Bufonidae: Rhinella) from Northern Peru. Taxonomy 2021, 1, 210-225. https://doi.org/10.3390/taxonomy1030015
AMA Style
Lehr E, Cusi JC, Rodriguez LO, Venegas PJ, García-Ayachi LA, Catenazzi A. A New Species of Toad (Anura: Bufonidae: Rhinella) from Northern Peru. Taxonomy. 2021; 1(3):210-225. https://doi.org/10.3390/taxonomy1030015
Chicago/Turabian StyleLehr, Edgar, Juan C. Cusi, Lily O. Rodriguez, Pablo J. Venegas, Luis A. García-Ayachi, and Alessandro Catenazzi. 2021. "A New Species of Toad (Anura: Bufonidae: Rhinella) from Northern Peru" Taxonomy 1, no. 3: 210-225. https://doi.org/10.3390/taxonomy1030015