Calypogeia (Calypogeiaceae, Marchantiophyta) in Pacific Asia: Updates from Molecular Revision with Particular Attention to the Genus in North Indochina

 , , , and
, , , and
Abstract
:1. Introduction
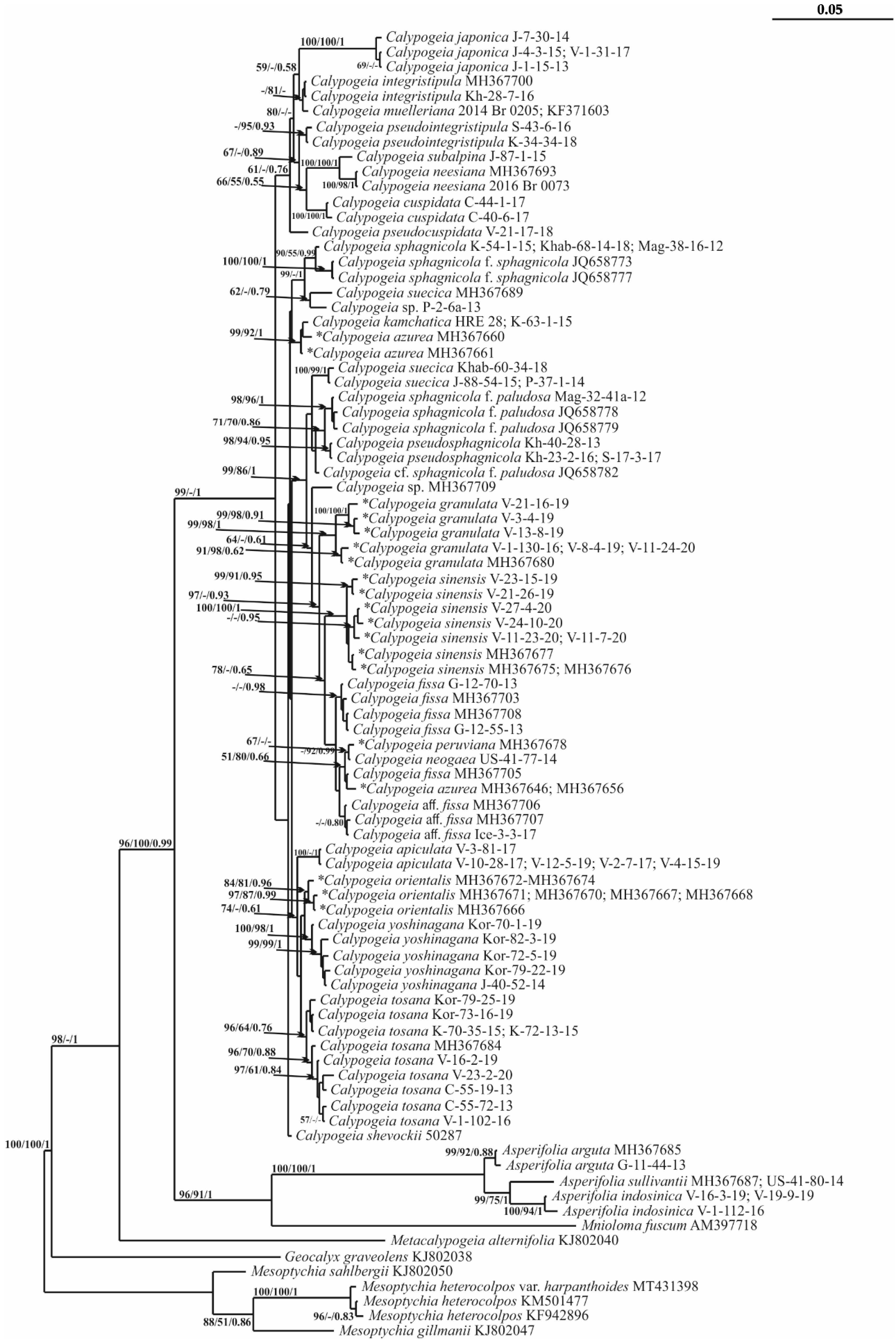
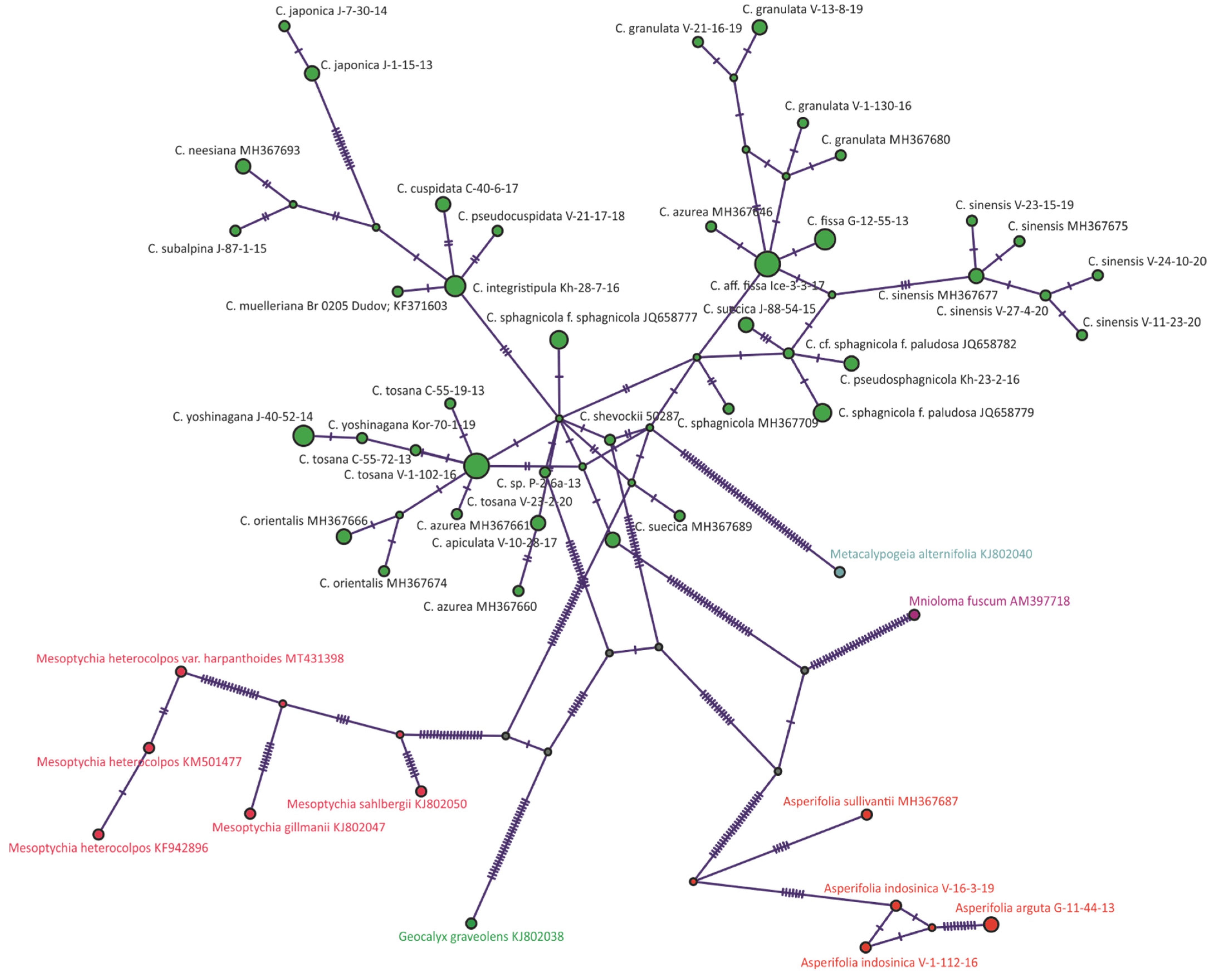
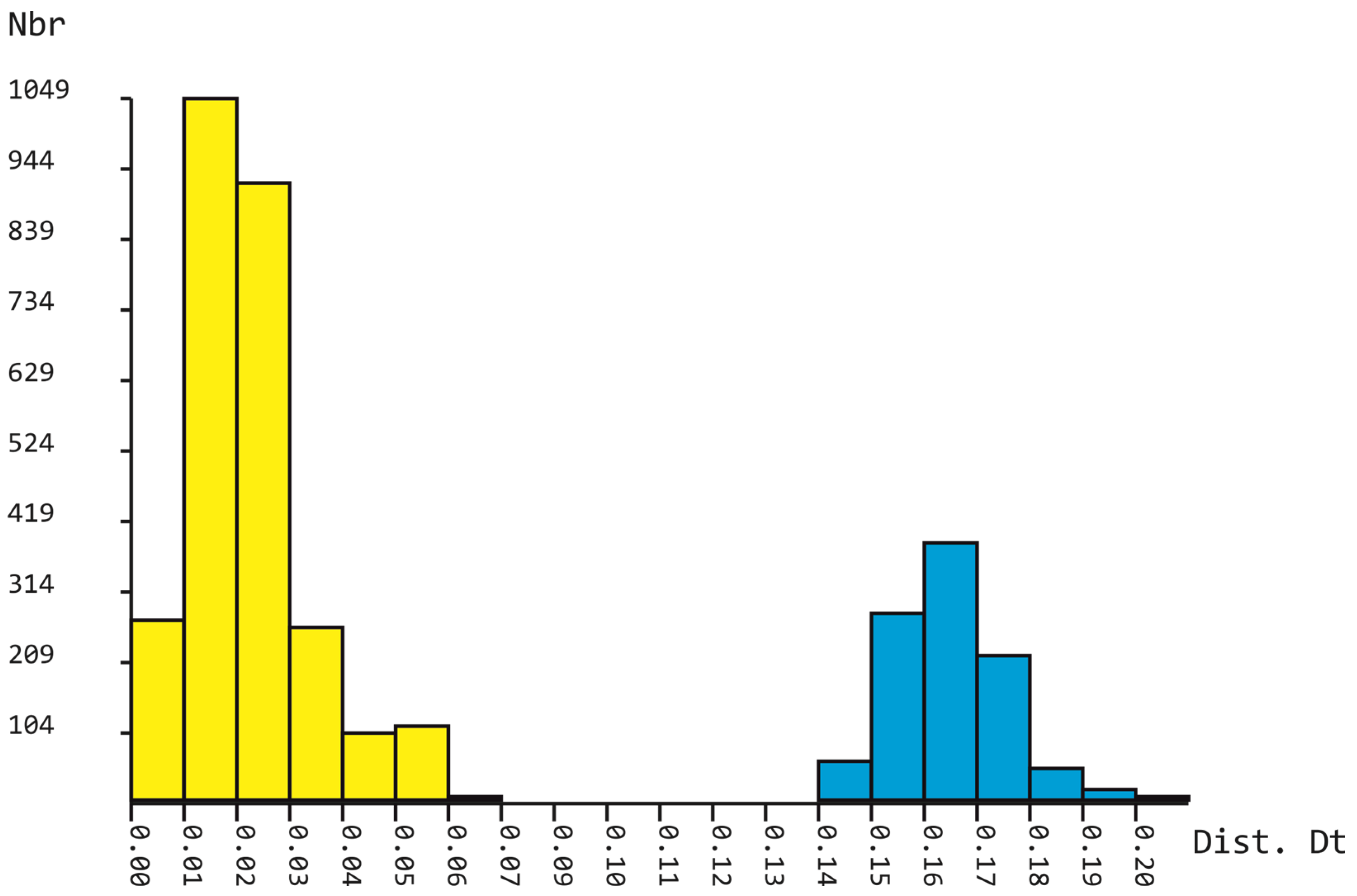
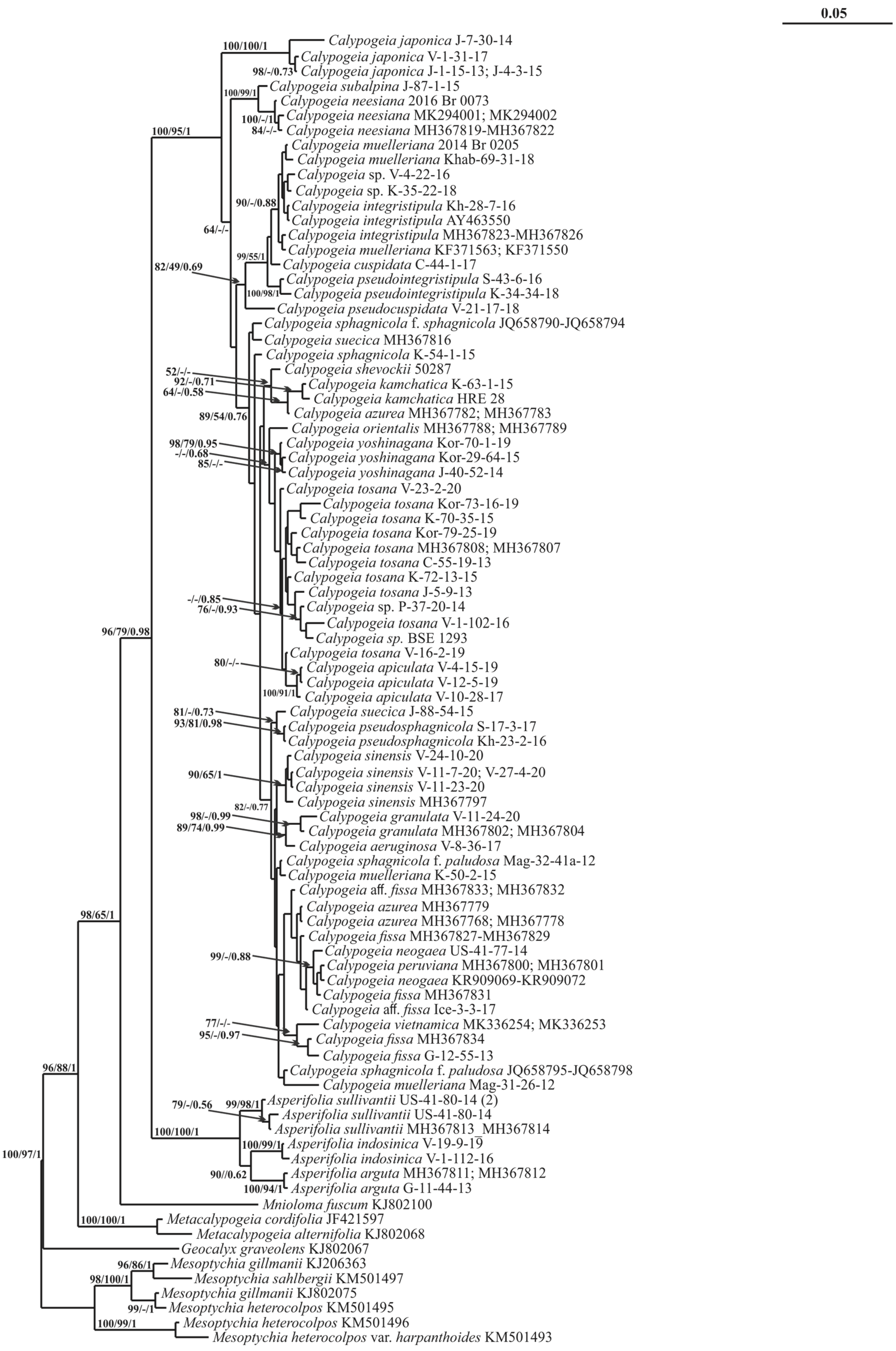
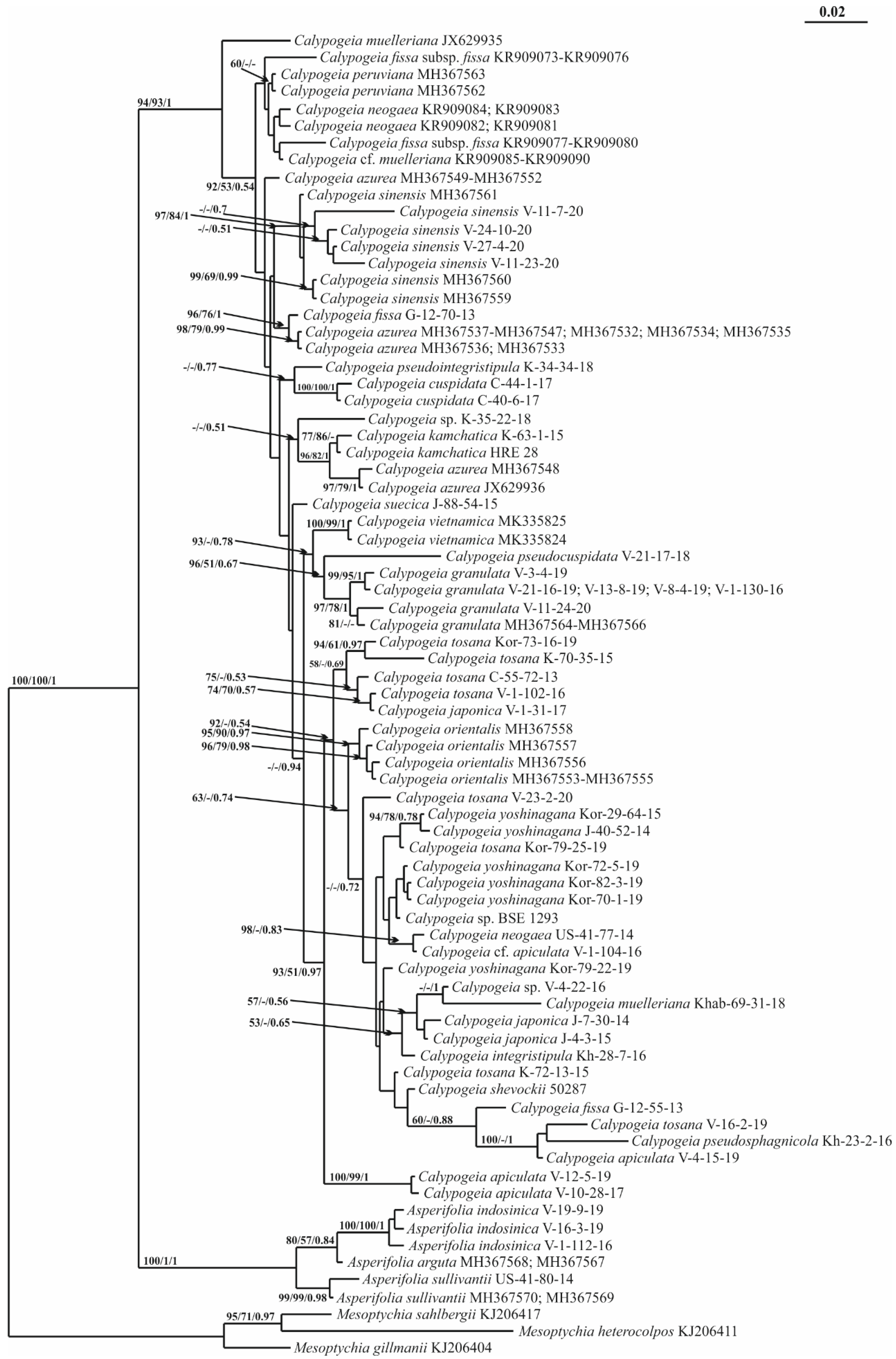
2. Results
3. Discussion
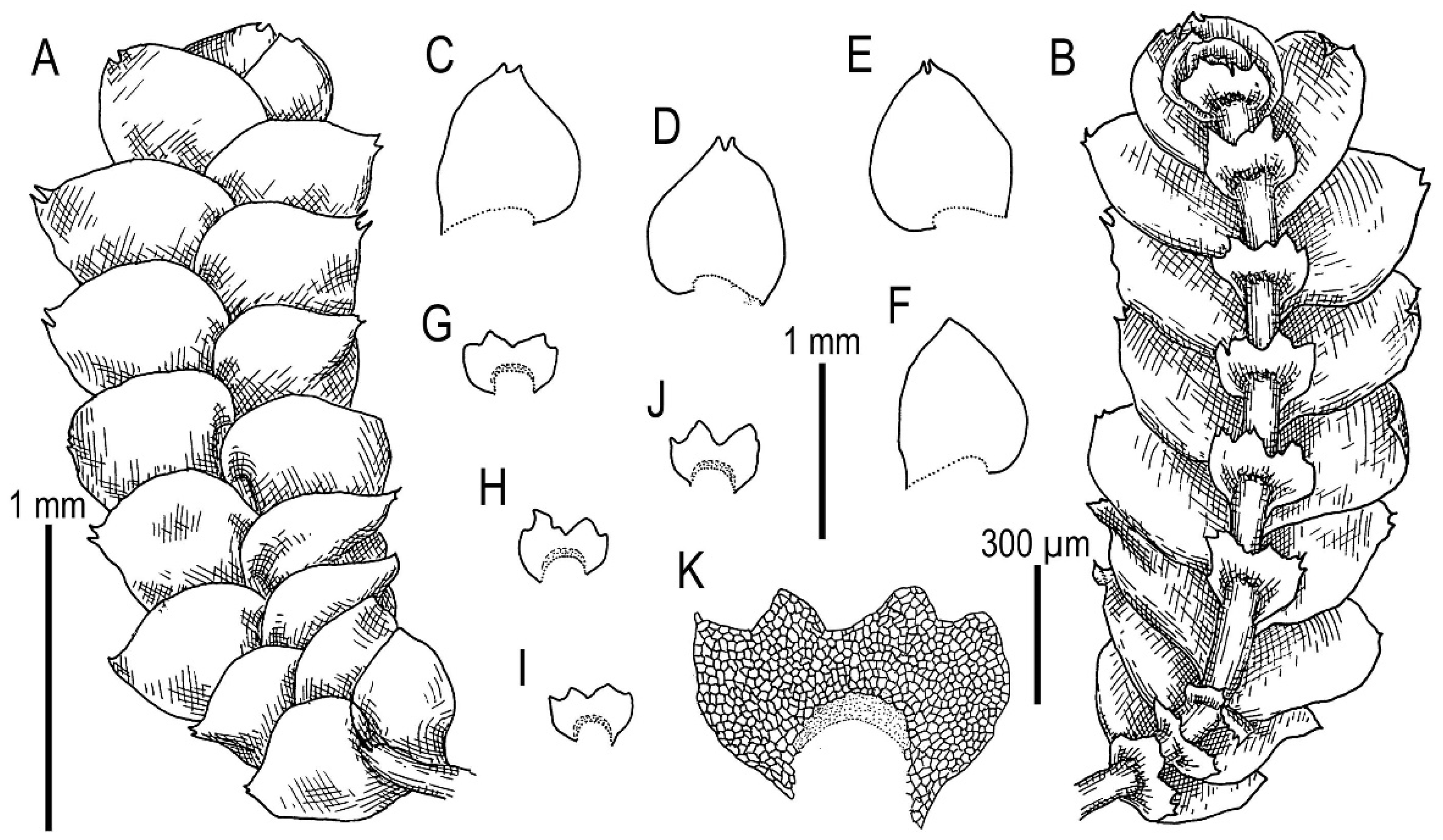
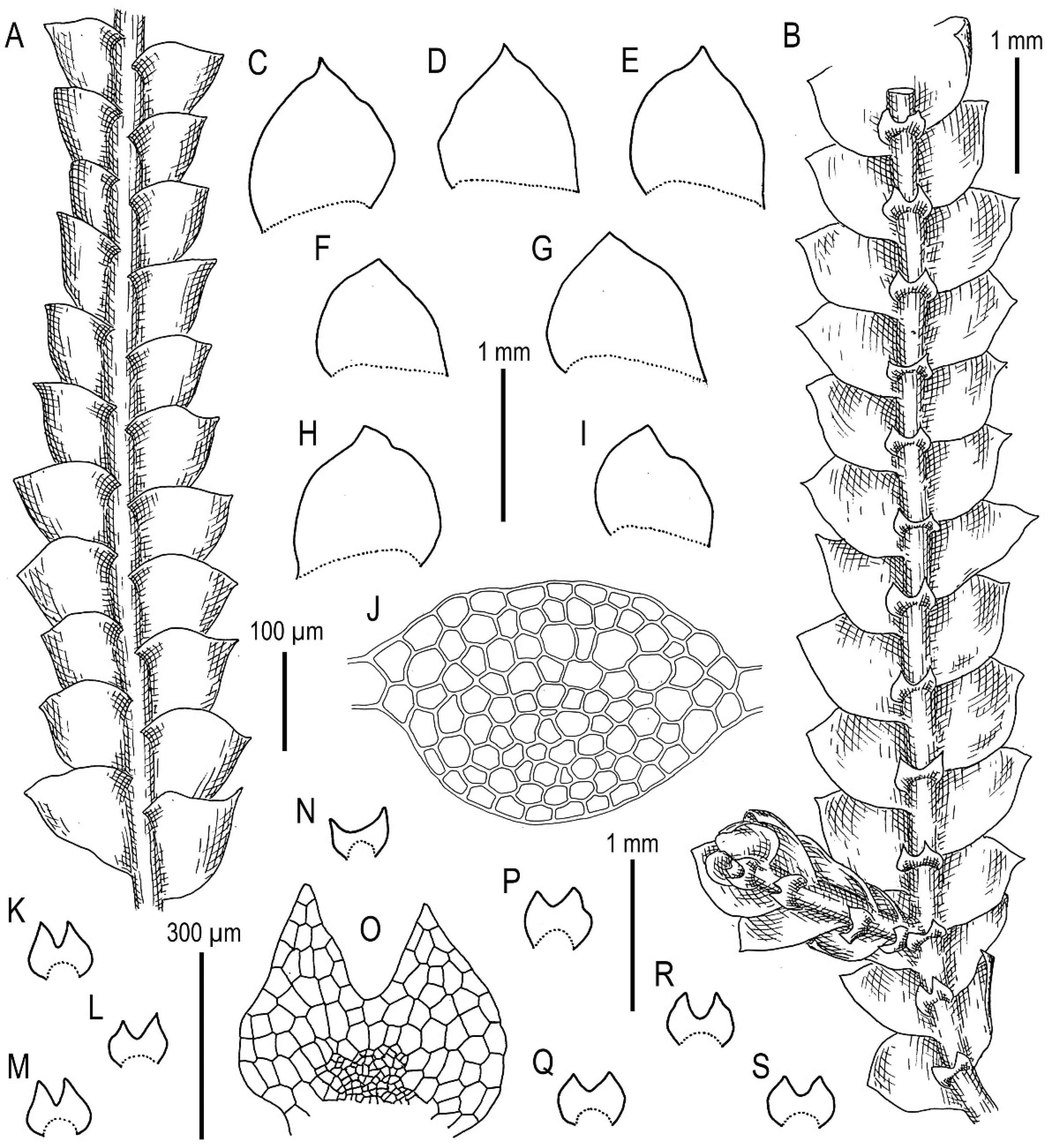
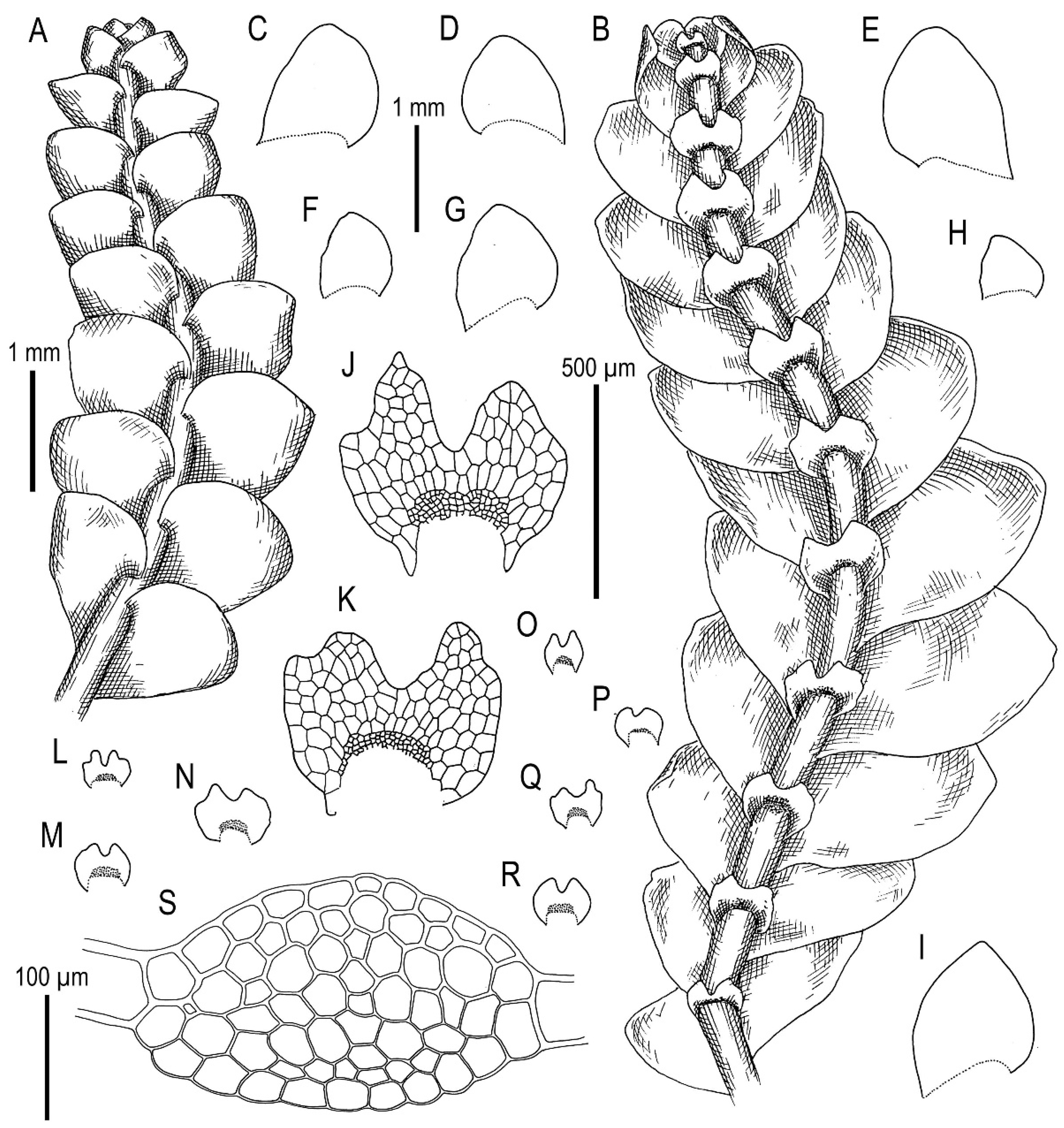
3.1. On Calypogeia arguta Nees et Mont. and Its Relatives
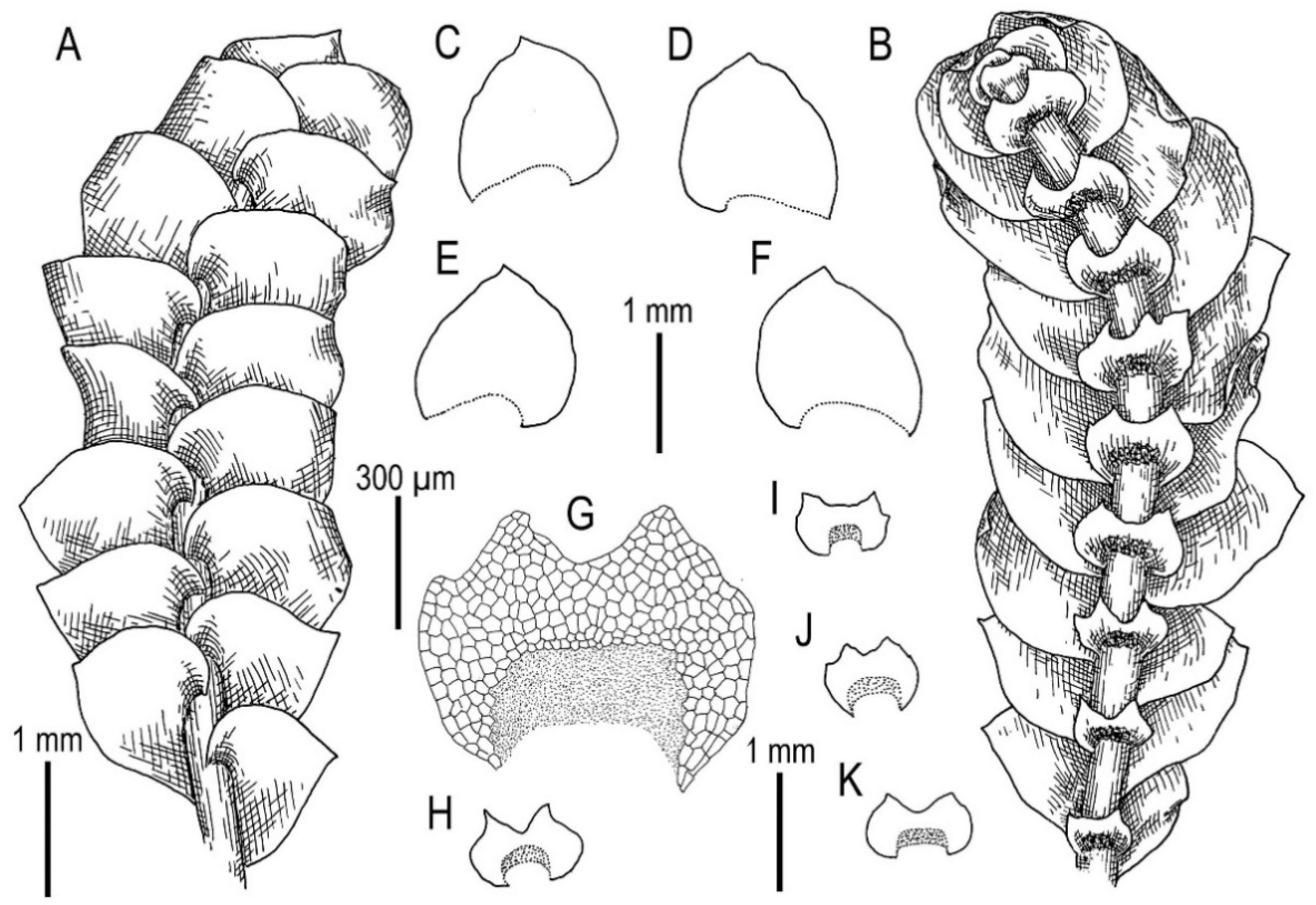
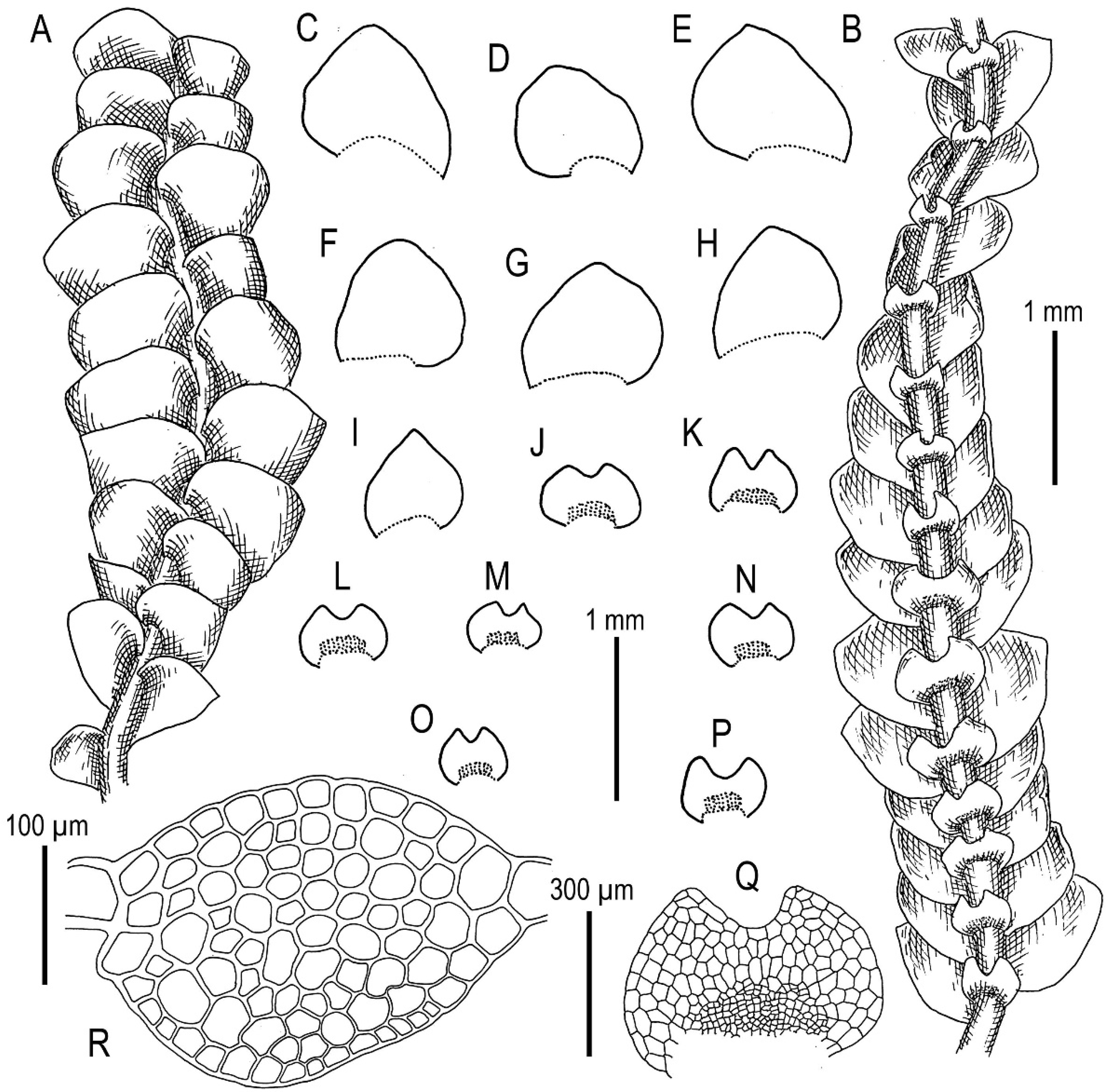
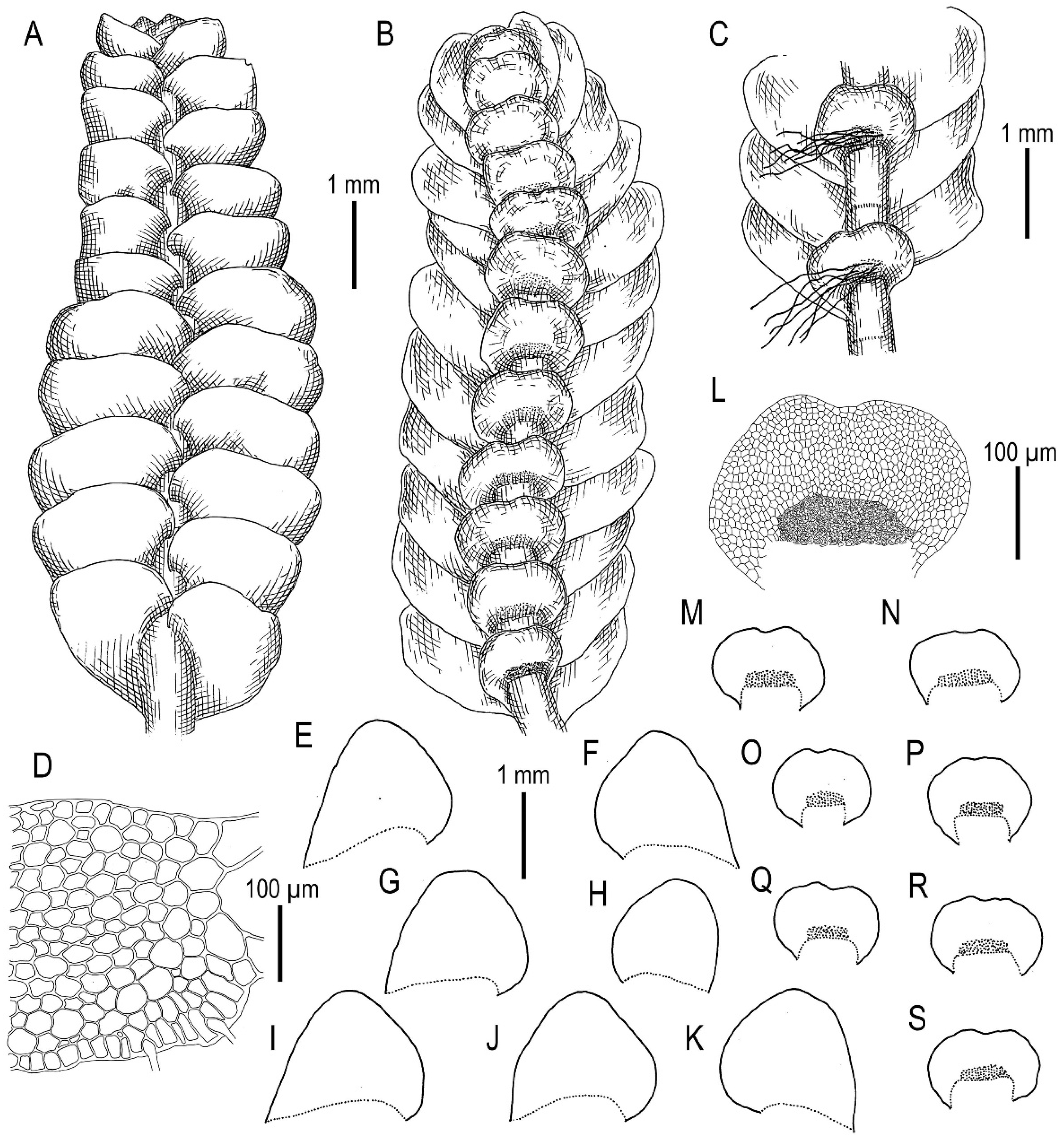
3.2. On Calypogeia pseudosphagnicola Bakalin, Troizk. et Maltseva
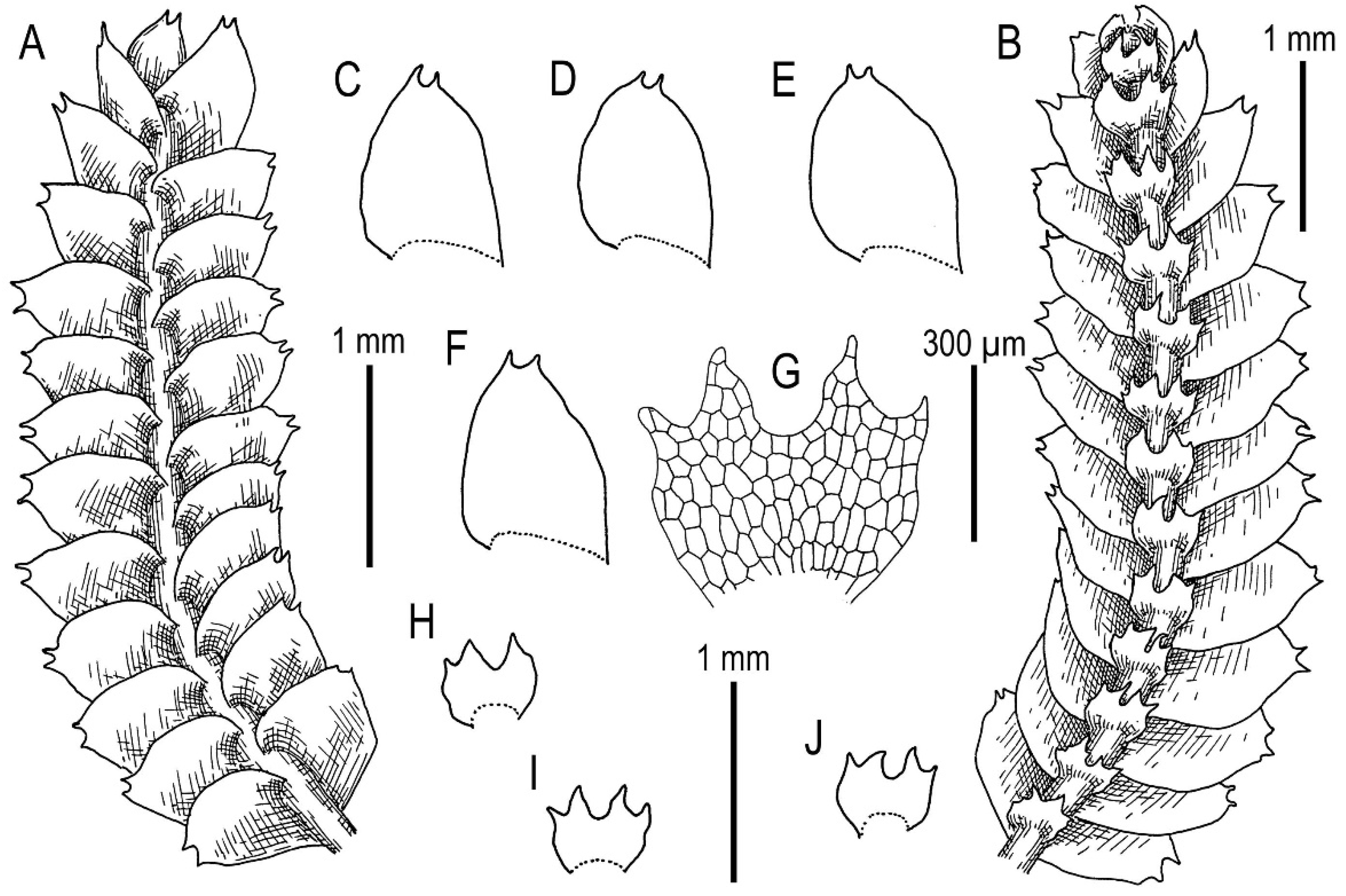
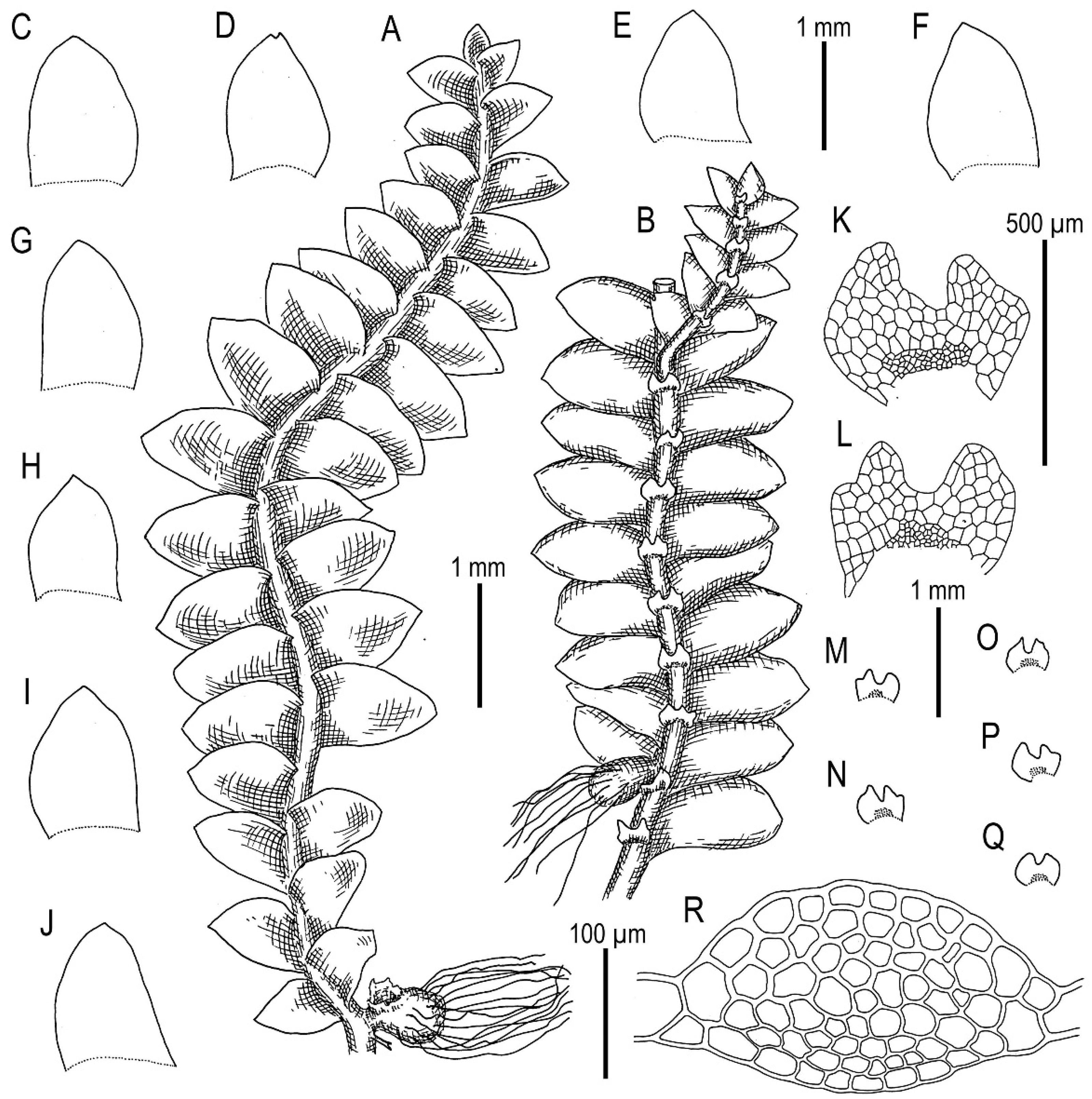
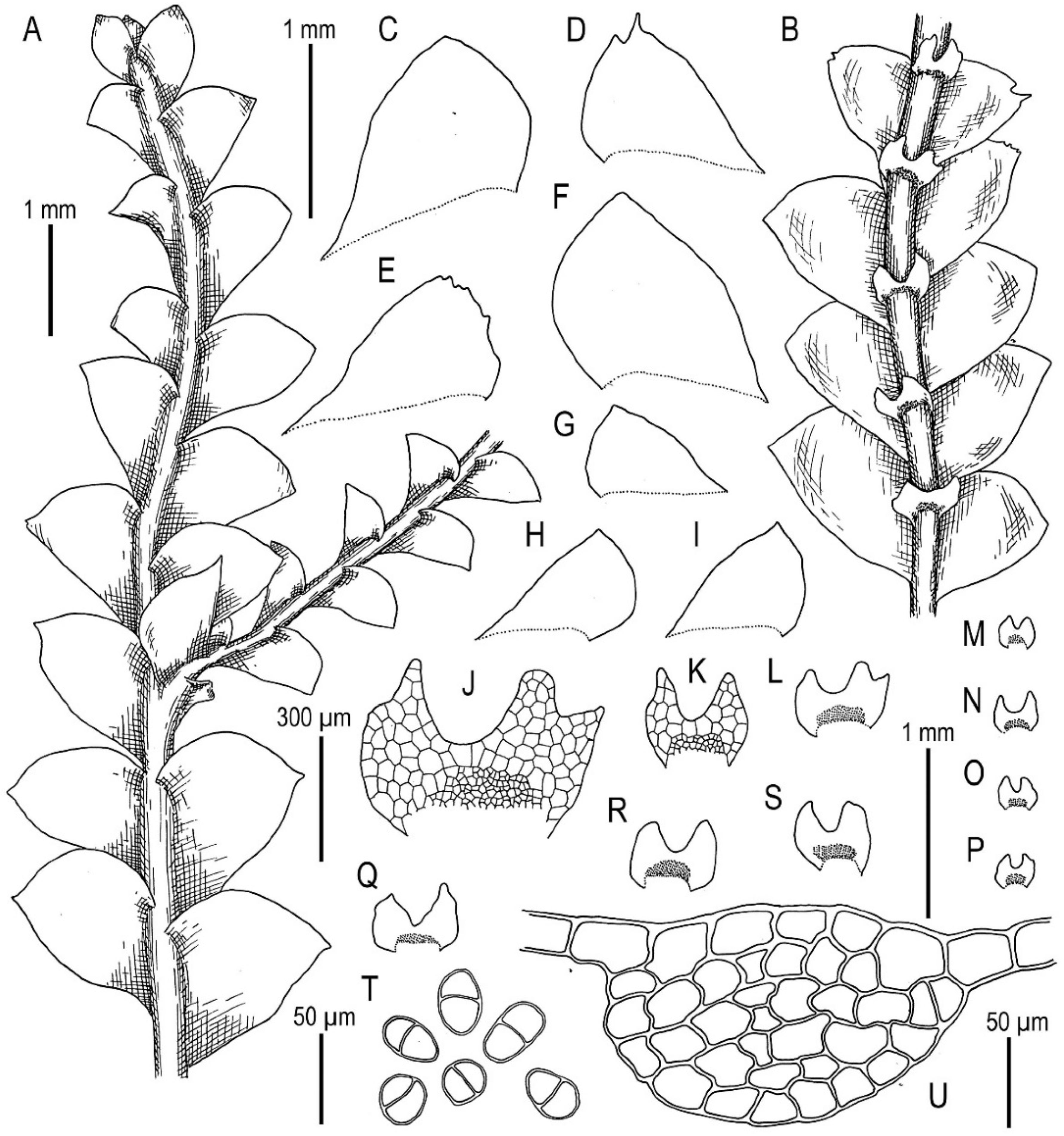
3.3. On Calypogeia subalpina Inoue
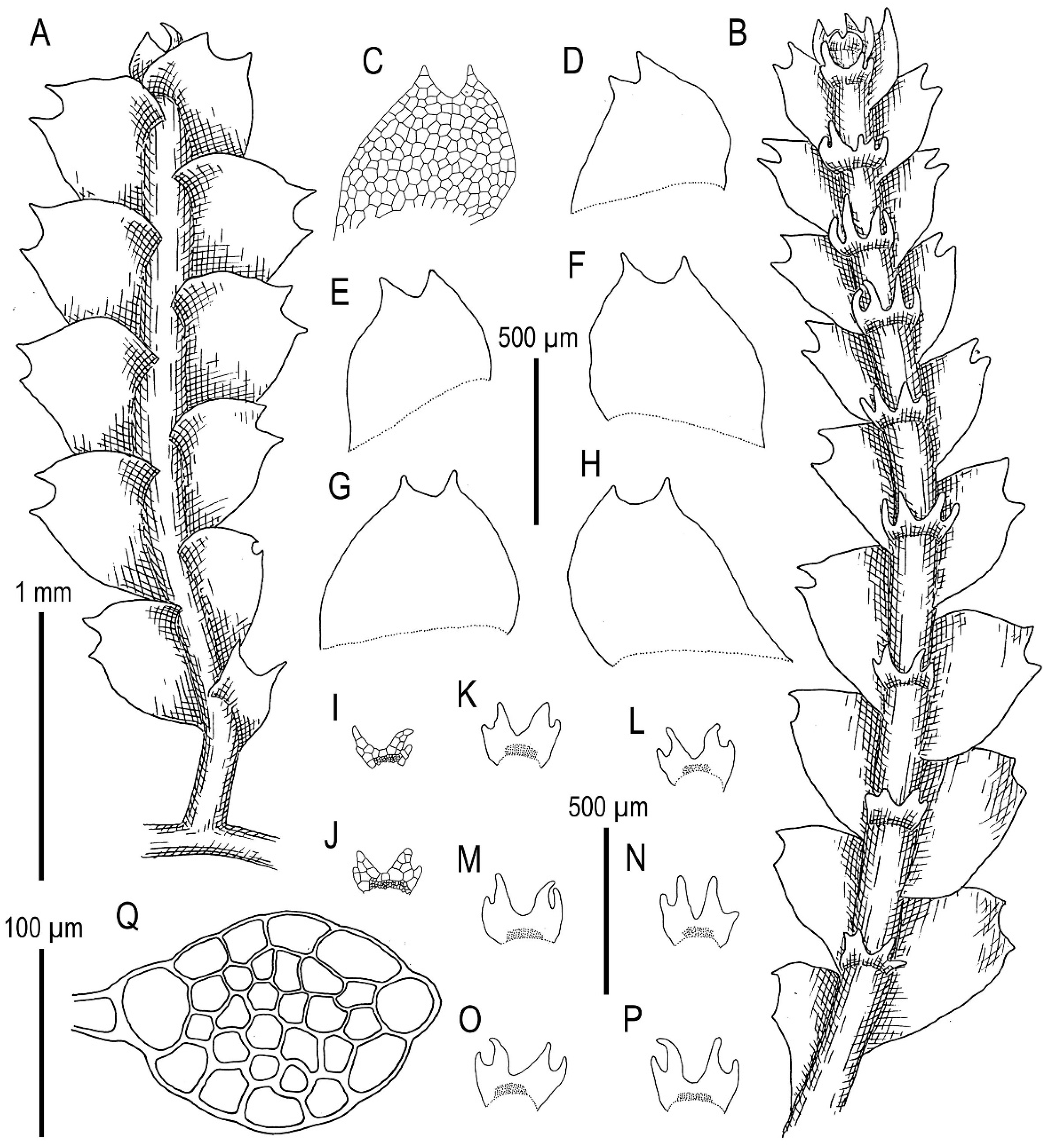
3.4. On the Complex Calypogeia neogaea (R.M. Schust.) Bakalin-C. kamchatica Bakalin, Troizk. et Maltseva-C. fissa (L.) Raddi
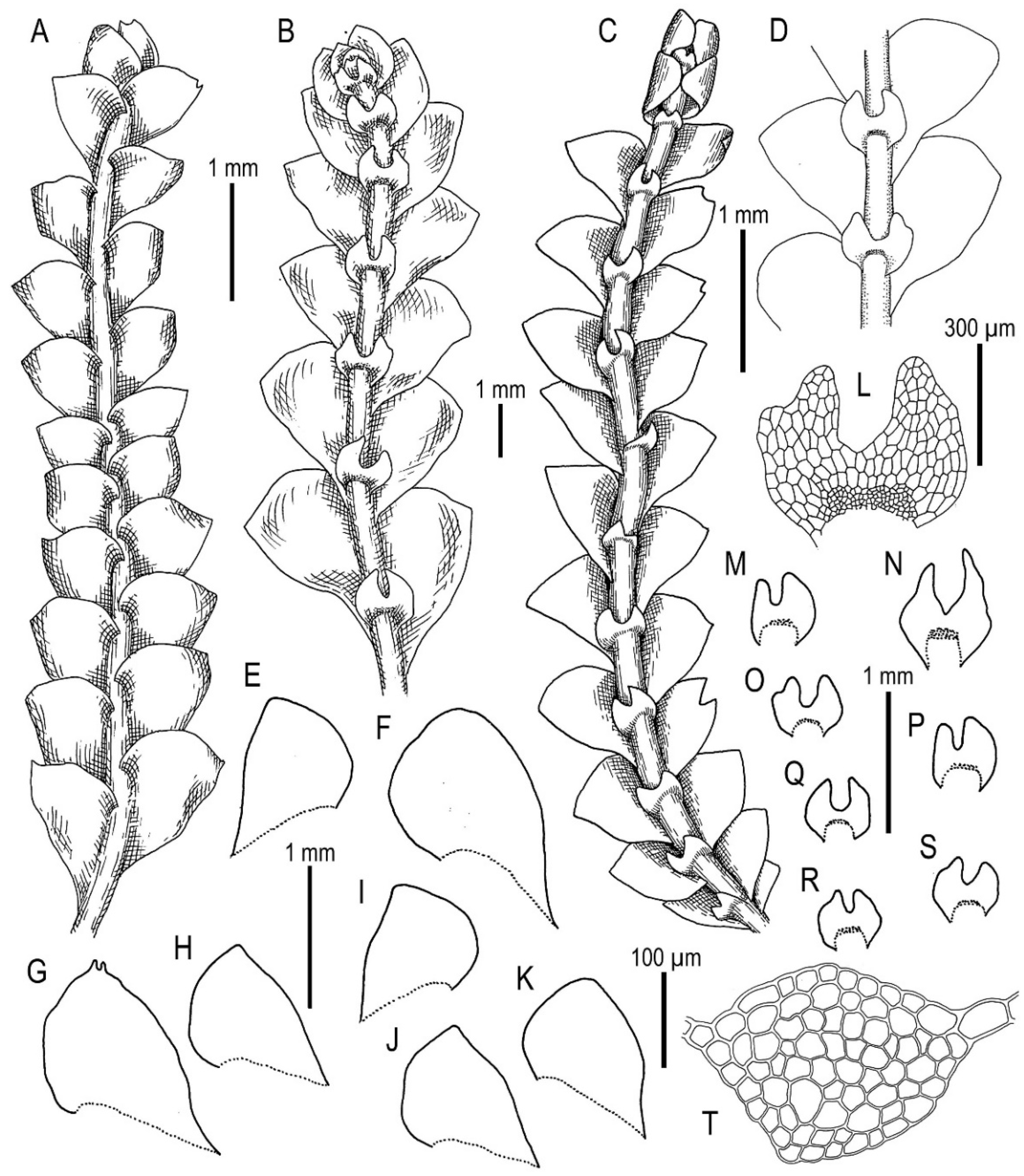
3.5. On Calypogeia tosana (Steph.) Steph. and C. yoshinagana Steph.
3.6. A New Calypogeia Species from California
3.7. On Calypogeia japonica Steph.
3.8. On Calypogeia pseudocuspidata Bakalin, Frank Müll. et Troizk.
3.9. On the Distribution of Calypogeia integristipula Steph. in the Pacific
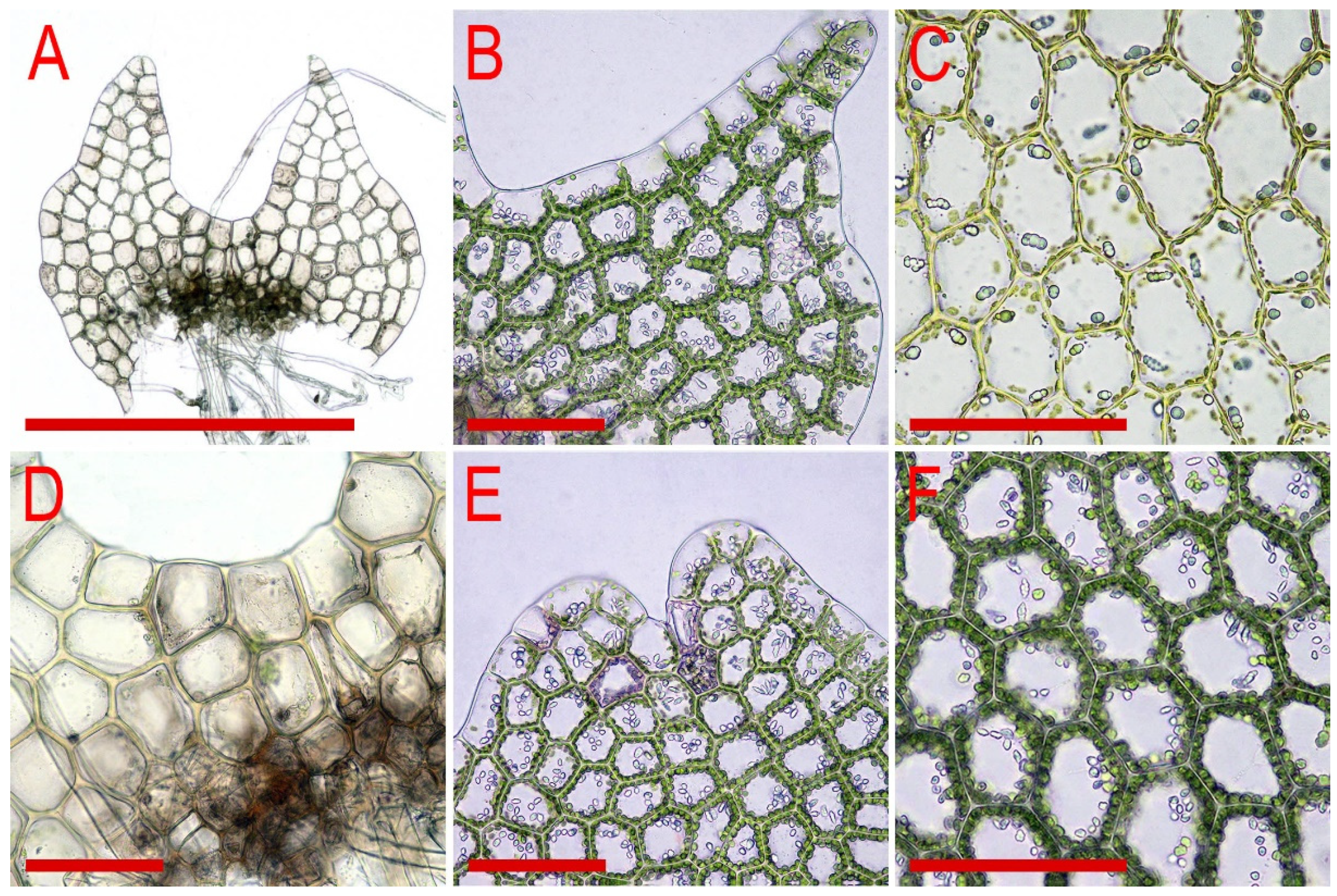
3.10. On Blue Oil-Bodied Calypogeia
3.11. On the Genus Calypogeia in Vietnam
3.11.1. Key to Calypogeia and Asperifolia taxa in Vietnam
3.11.2. Annotated List of Calypogeia and Asperifolia taxa in Vietnam
3.12. On the Genus Calypogeia in Myanmar
4. Taxonomic Treatment
5. Materials and Methods
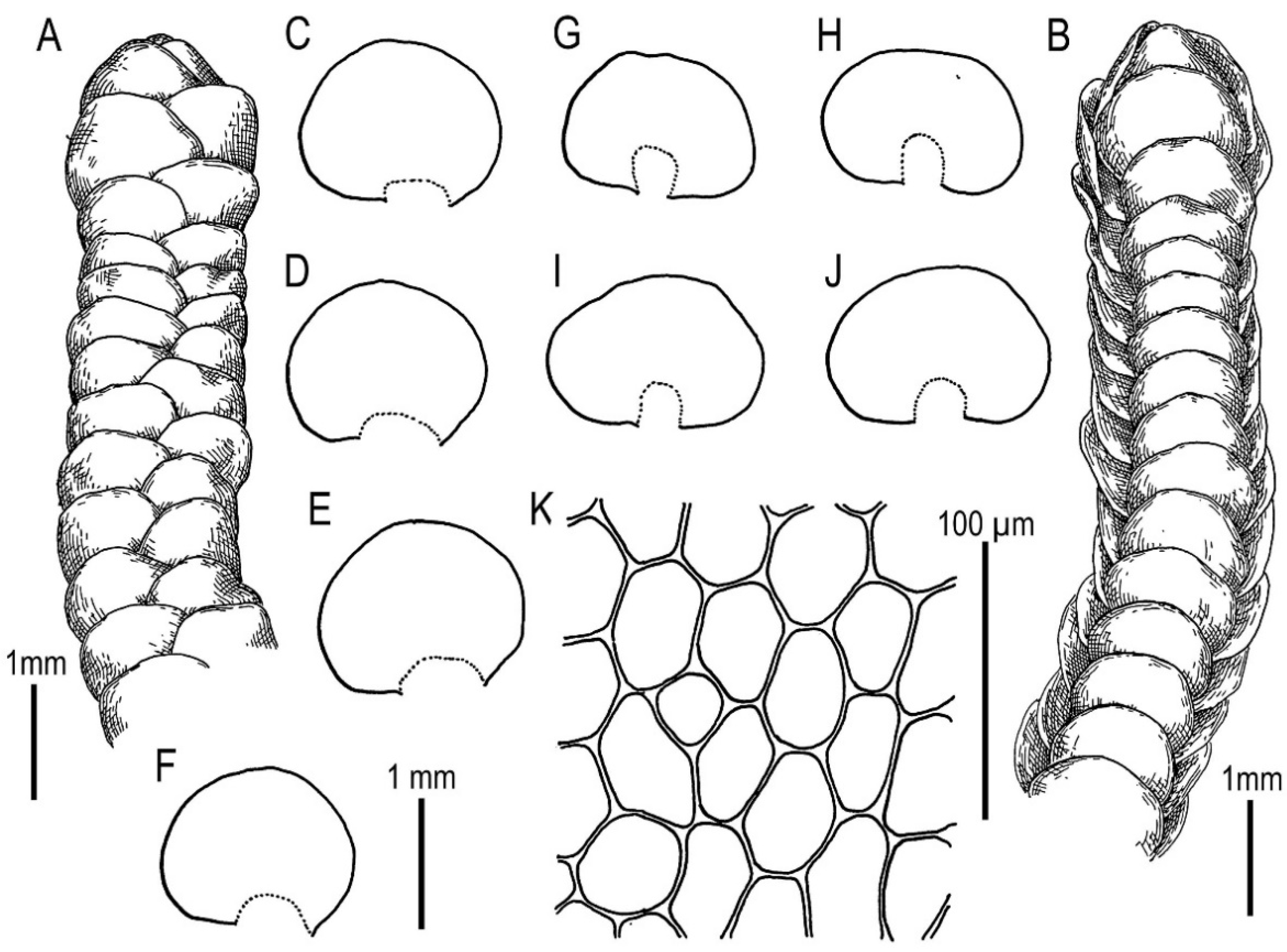
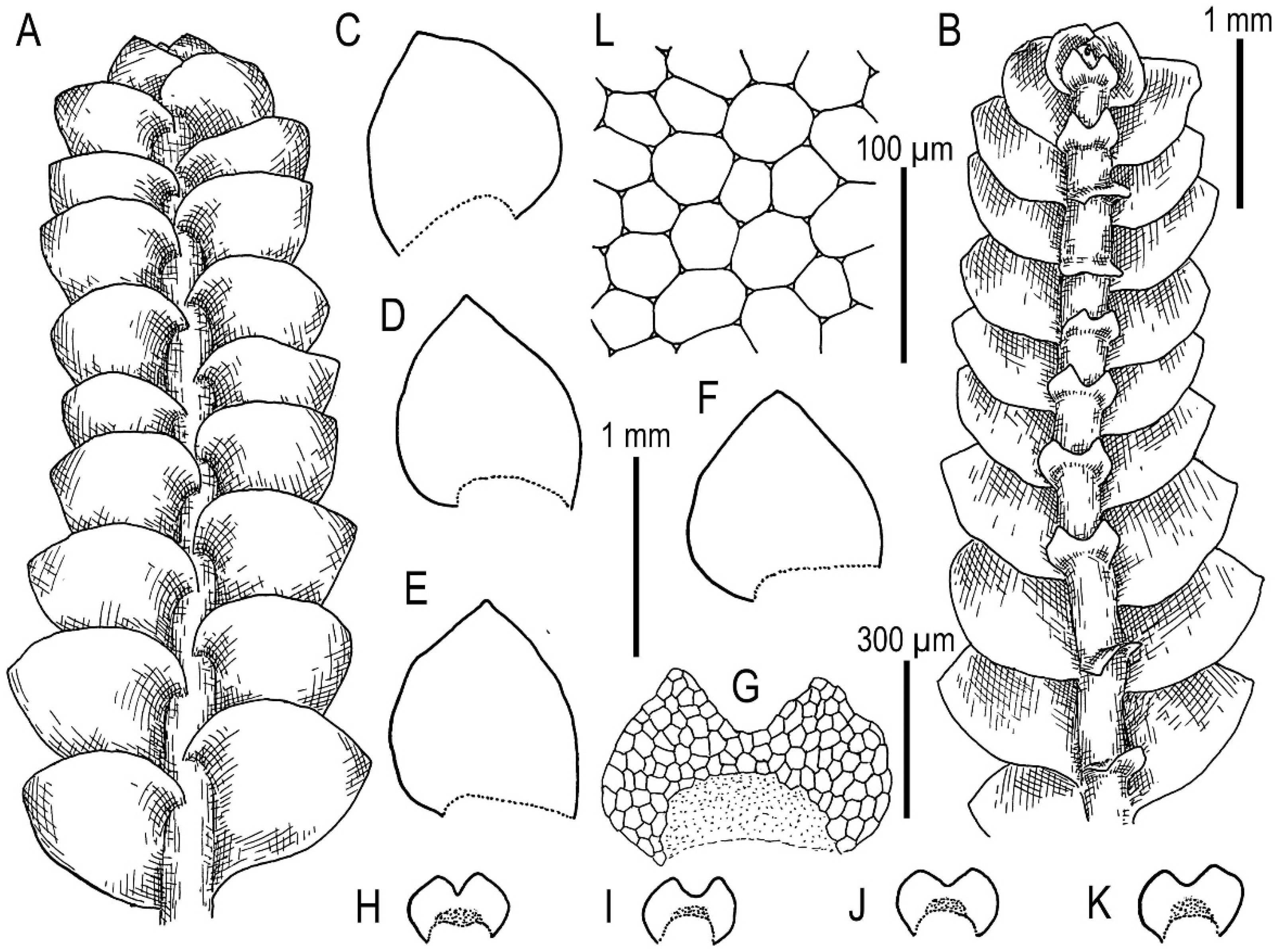
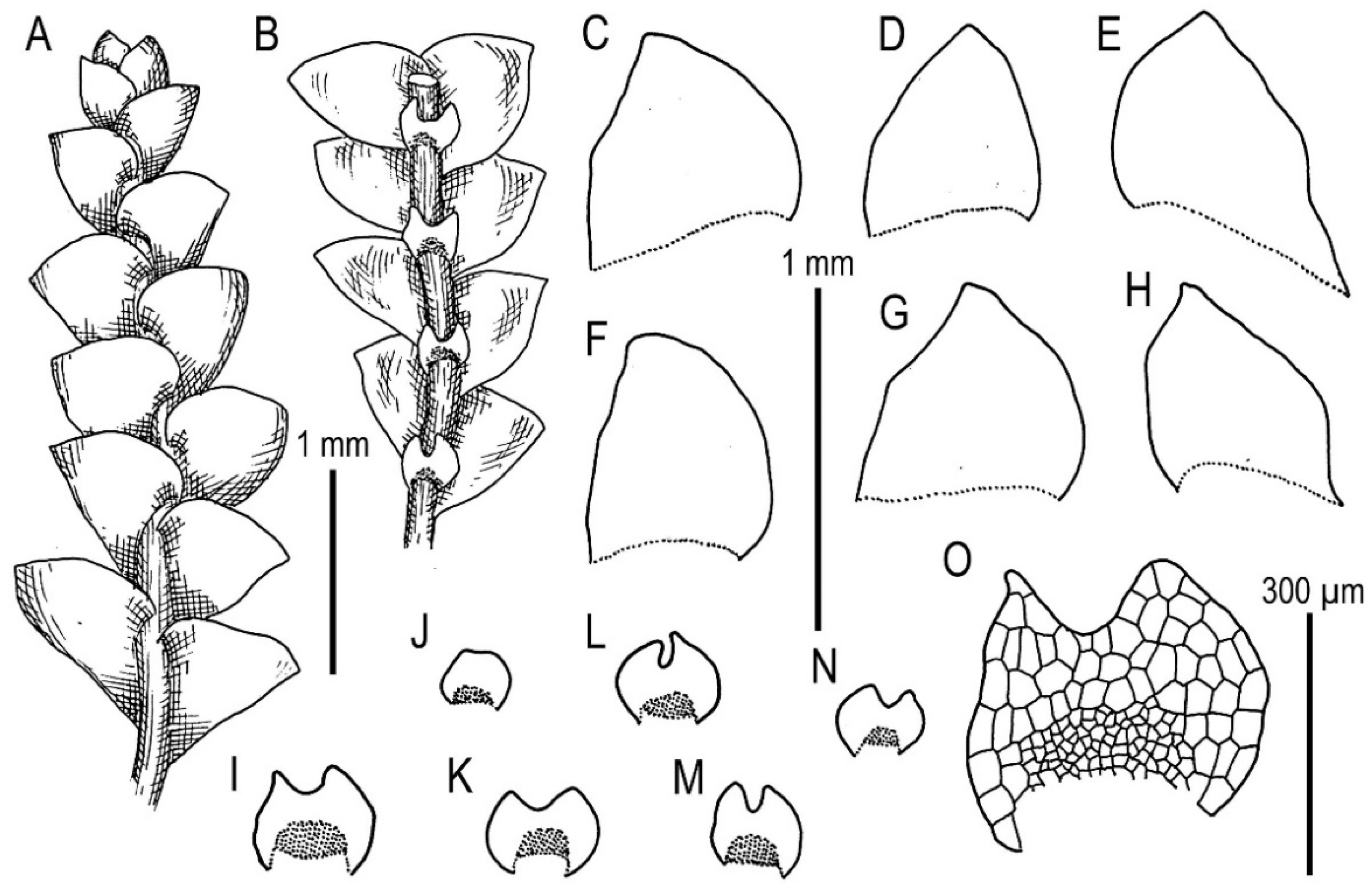
5.1. Morphology Comparison
5.2. DNA Isolation, Amplification, and Sequencing
5.3. Phylogenetic Analyses
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bakalin, V.A.; Klimova, K.G.; Nguyen, V.S. A review of Calypogeia (Marchantiophyta) in the eastern Sino-Himalaya and Meta-Himalaya based mostly on types. PhytoKeys 2020, 153, 111–154. Available online: https://phytokeys.pensoft.net/article/52920/ (accessed on 15 February 2022). [CrossRef] [PubMed]
- Bakalin, V.; Vilnet, A.; Klimova, K.; Nguyen, V.S. Calypogeia vietnamica sp. nov. (Calypogeiaceae, Hepaticae) from North Vietnam and diversification in Calypogeia taxa with blue oil bodies. Herzogia 2019, 32, 219–229. [Google Scholar] [CrossRef]
- Buczkowska, K. Genetic differentiation of Calypogeia fissa Raddi (Hepaticae, Jungermanniales) in Poland. Plant. Syst. Evol. 2004, 247, 187–201. [Google Scholar] [CrossRef]
- Buczkowska, K. The genus Calypogeia Raddi (Jungermanniales, Hepaticae) in Poland, biometrical analysis of morphological and anatomical variation. Nova Hedwig. 2004, 78, 121–146. [Google Scholar] [CrossRef]
- Buczkowska, K.; Bączkiewicz, A. New taxon of the genus Calypogeia (Jungermanniales, Hepaticae) in Poland. Acta Soc. Bot. Pol. 2011, 80, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Buczkowska, K.; Gonera, P.; Bączkiewicz, A.; Rosadziński, S.; Rabska, M. Chloroplast DNA sequences confirmed genetic divergence within Calypogeia muelleriana (Calypogeiaceae, Marchantiophyta). Biodivers. Res. Conserv. 2015, 32, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Buczkowska, K.; Bakalin, V.; Bączkiewicz, A.; Aguero, B.; Gonera, P.; Ślipiko, M.; Szczecińska, M.; Sawicki, J. Does Calypogeia azurea (Calypogeiaceae, Marchantiophyta) occur outside Europe? Molecular and morphological evidence. PLoS ONE 2018, 13, e0204561. [Google Scholar] [CrossRef]
- Warnstorf, C. Die europäischen Artengruppen der Gattung Calypogeia Raddi (1820). Bryol. Z. 1917, 1, 97–114. [Google Scholar]
- Schuster, R.M. The Hepaticae and Anthocerotae of North America. II; Columbia University Press: New York, NY, USA, 1969; 1062p. [Google Scholar]
- Müller, K. Die Lebermoose Europas. In Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Österreich und der Schweiz, 3rd ed.; Akademische Verlagsgesellschaft: Leipzig, Germany, 1956; Volume 6, pp. 917–1076. [Google Scholar]
- Inoue, H. Miscellaneous notes on hepatics of Japan (3). Jpn. J. Bot. 1962, 37, 101–105. [Google Scholar]
- Inoue, H. Notes on the Calypogeia neesiana complex in Japan. Mem. Natl. Sci. Mus. 1971, 4, 55–58. [Google Scholar]
- Konstantinova, N.A.; Bakalin, V.A.; Andrejeva, E.N.; Bezgodov, A.G.; Borovichev, E.A.; Dulin, M.V.; Mamontov, Y.S. Checklist of liverworts (Marchantiophyta) of Russia. Arctoa 2009, 18, 1–64. [Google Scholar] [CrossRef]
- Bakalin, V.A. Liverworts of Kamchatka: The results of study. In Proceedings of the VII International Scientific Conference “Conservation of Biodiversity of Kamchatka and Coastal Waters”, Petropavlovsk-Kamchatsky, Russia, 28–29 November 2007; pp. 6–14. [Google Scholar]
- Bakalin, V.A. Distribution of Bryophytes in the Russian Far East. Part. I; Hepatics; Izd-vo DVFU: Vladivostok, Russia, 2010; 175p. [Google Scholar]
- Stotler, R.E.; Crandall-Stotler, B. A Synopsis of the Liverwort Flora of North America North of Mexico. Ann. Mo. Bot. Gard. 2017, 102, 574–709. [Google Scholar] [CrossRef]
- Söderström, L.; Váňa, J.; Hagborg, A.; von Konrat, M. Notes on Early Land Plants Today. 35. Notes on Lophoziaceae (Marchantiophyta). Phytotaxa 2013, 97, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Hattori, S. Hepaticae and Anthocerotae of Mt. Chokai, Northern Japan. J. Hattori Bot. Lab. 1966, 29, 267–278. [Google Scholar]
- Choi, S.S.; Bakalin, V.; Park, S.J. Integrating continental mainland and islands in temperate East Asia: Liverworts and hornworts of the Korean Peninsula. PhytoKeys 2021, 176, 131–226. [Google Scholar] [CrossRef] [PubMed]
- Stephani, F. Species hepaticarum 3. Bull. L’herbier Boissier (Sér. 2) 1908, 8, 661–696. [Google Scholar] [CrossRef]
- Bonner, C.E.B. Index Hepaticarum; J. Cramer: Weinheim, Germany, 1963; pp. 3–4, 1–926. [Google Scholar]
- Grolle, R. Verzeichnis der Lebermoose Europas und benachbarter Gebiete. Feddes Repert. 1976, 87, 171–279. [Google Scholar] [CrossRef]
- Meuche, D.; Huneck, S. Azulene aus Calypogeia trichomanis (L.) Corda. Chem. Ber. 1966, 99, 2669–2674. [Google Scholar] [CrossRef]
- Bakalin, V.; Nguyen, V.S.; Klimova, K.; Bakalin, D.; Nguyen, H.M. Where and how many new additions to the liverwort flora of Vietnam may be found? Bryologist 2021, 124, 391–402. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Nguyen, V.S.; Borovichev, E.A. New liverwort records for Vietnam. J. Bryol. 2018, 40, 68–73. [Google Scholar] [CrossRef]
- Shu, L.; Xiang, Y.-L.; Cheng, X.-F.; Wei, Y.-M.; Wang, J.; Zhang, L.-N.; Li, W.; Yin, X.-B.; Zhang, W.-P.; Zhao, C.-X.; et al. New Liverwort and Hornwort Records for Vietnam. Cryptogam. Bryol. 2017, 38, 411–445. [Google Scholar] [CrossRef]
- Stephani, F. Hepaticae sandvicenses. Bull. L’herbier Boissier 1897, 5, 840–849. [Google Scholar]
- Yang, J.D.; Lin, S.H. Calypogeia aeruginosa Mitten, a newly recorded liverwort to Taiwan. Endem. Species Res. 2009, 11, 93–99. [Google Scholar]
- Inoue, H. A new species of Calypogeia Raddi. Jpn. J. Bot. 1968, 43, 468–472. [Google Scholar]
- Furuki, T.; Ota, M. Taxonomical study of Calypogeia japonica (Hepaticae) described from Japan. Bryol. Res. 2001, 7, 381–384. [Google Scholar]
- Yao, K.Y.; Yang, J.D. Calypogeia japonica Stephani, a Newly Recorded Liverwort in Taiwan. Guójiā Gōngyuán Xuébào Èr (Natl. Park J.) 2015, 25, 76–80. [Google Scholar]
- Inoue, H. Illustrations of Japanese Hepaticae; Tsukiji Shokan Publishing Company: Tsukuba, Japan, 1974; 189p. [Google Scholar]
- Milyutina, I.A.; Goryunov, D.V.; Ignatov, M.S.; Ignatova, E.A.; Troitsky, A.V. The phylogeny of Schistidium (Bryophyta, Grimmiaceae) based on the primary and secondary structure of nuclear rDNA internal transcribed spacers. Mol. Biol. 2010, 44, 883–897. [Google Scholar] [CrossRef]
- Groth, H.; Helms, G.; Heinrichs, J. The systematic status of Plagiochila sects. Bidentes Carl and Caducilobae Inoue (Hepaticae) inferred from nrDNA ITS sequences. Taxon 2003, 51, 675–684. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Snisky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Guy Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Bakalin, V.; Maltseva, Y.; Vilnet, A.; Choi, S.S. The transfer of Tritomaria koreana to Lophozia has led to recircumscription of the genus and shown convergence in Lophoziaceae (Hepaticae). Phytotaxa 2021, 512, 41–56. [Google Scholar] [CrossRef]
- Pacak, A.; Szweykowska-Kulinska, Z. Molecular data concerning alloploid character and the origin of chloroplast and mitochondrial genomes in the liverwort species Pellia borealis. J. Plant Biotechnol. 2000, 2, 101–108. [Google Scholar]
- Katoh, K.; Standley, D.N. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van Der Mark, P.; Ayres, D.L.; Darling, S.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K. TCS: Estimating gene genealogies. In Proceedings of the 16th International Parallel and Distributed Processing Symposium, Fort Lauderdale, FL, USA, 15–19 April 2002; p. 184. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. Popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Species Name | Accepted Name | Label | GenBank Accession Number | ||
|---|---|---|---|---|---|
| ITS | trnL | trnG | |||
| Calypogeia aeruginosa Mitt. | Calypogeia aeruginosa Mitt. | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-8-36-17 (VBGI) | - | OL408961 | - |
| Calypogeia angusta Steph. | Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Japan, Honshu Island, V.A. Bakalin, J-88-54-15 (VBGI) | OL689055 | OL408989 | OL311475 |
| Calypogeia apiculata (Steph.) Steph. | Calypogeia apiculata (Steph.) Steph. | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-2-7-17 (VBGI) | - | - | OL311465 |
| Calypogeia apiculata (Steph.) Steph. | Calypogeia apiculata (Steph.) Steph. | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-3-81-17 (VBGI) | - | - | OL311466 |
| Calypogeia apiculata (Steph.) Steph. | Calypogeia apiculata (Steph.) Steph. | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-10-28-17 (VBGI) | OL689064 | OL409000 | OL311463 |
| Calypogeia arguta Nees et Mont. | Asperifolia arguta (Nees et Mont.) Troizk., Bakalin, Maltseva | Germany, A. Solga & D. Quandt, POZW 39075 | MH367567 | MH367811 | MH367685 |
| Calypogeia arguta Nees et Mont. | Asperifolia arguta (Nees et Mont.) Troizk., Bakalin, Maltseva | Spain, Galicia, A. Schäfer-Verwimp, S-V 31365 | MH367568 | MH367812 | * MH367686 |
| Calypogeia arguta Nees et Mont. | Asperifolia indosinica Troizk. et Bakalin | Vietnam, Lai Châu Province, V.A. Bakalin & K.G. Klimova, V-16-3-19 (VBGI) | OL689056 | OL408990 | OL311426 |
| Calypogeia arguta Nees et Mont. | Asperifolia indosinica Troizk. et Bakalin | Vietnam, Sơn La Province, V.A. Bakalin & K.G. Klimova, V-19-9-19 (VBGI) | OL689065 | OL409001 | OL311427 |
| Calypogeia arguta Nees et Mont. | Asperifolia arguta (Nees et Mont.) Troizk., Bakalin, Maltseva | Georgia, Adjaria, V.A. Bakalin, G-11-44-13 (VBGI) | OL689057 | OL409010 | OL311472 |
| Calypogeia arguta Nees et Mont. | Asperifolia indosinica Troizk. et Bakalin | Vietnam, Lao Cai Province, V.A. Bakalin, V-1-112-16 (VBGI) | OL689079 | OL409012 | OL311470 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | S Poland, K. Buczkowska & A. Baczkiewicz, POZW 41776 | MH367534 | MH367768 | MH367646 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | Germany, A. Schäfer-Verwimp, S-V 31563 | MH367544 | MH367778 | MH367656 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | Germany, A. Schäfer-Verwimp, S-V 30124 | MH367545 | MH367779 | * MH367657 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | North America, Canada, British Columbia, B. Aguero, POZW 42447 | MH367548 | MH367782 | MH367660 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | Russia, Buryatiya Republic, N. Konstantinova & A. Savchenko, 20-01 (KPABG) | JX629936 | * JX630063 | - |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | NE Poland, K. Buczkowska & A. Bączkiewicz, POZW 41778 | MH367532 | * MH367766 | * MH367644 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | S Poland, A. Bączkiewicz & K. Buczkowska, POZW 41390 | MH367533 | * MH367767 | * MH367645 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | S Poland, K. Buczkowska & A. Bączkiewicz, POZW 41371 | MH367535 | * MH367769 | * MH367647 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | S Poland, K. Buczkowska & A. Bączkiewicz, POZW 41378 | MH367536 | * MH367770 | * MH367648 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | SE Poland, K. Buczkowska & B. Chmielewska, POZW 41925 | MH367537 | * MH367771 | * MH367649 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | SE Poland, K. Buczkowska & B. Chmielewska, POZW 41929 | MH367538 | * MH367772 | * MH367650 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | SE Poland, K. Buczkowska & B. Chmielewska, POZW 41948 | MH367539 | * MH367773 | * MH367651 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | SE Poland, K. Buczkowska & B. Chmielewska, POZW 42390 | MH367540 | * MH367774 | * MH367652 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | SE Poland, K. Buczkowska & B. Chmielewska, POZW 42373 | MH367541 | * MH367775 | * MH367653 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | SE Poland, K. Buczkowska & B. Chmielewska, POZW 42360 | MH367542 | * MH367776 | * MH367654 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | Austria, A. Schäfer-Verwimp, S-V 27519/A | MH367543 | * MH367777 | * MH367655 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | Romania, S. Ştefănut, POZW 42609 | MH367546 | * MH367780 | * MH367658 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | Russia, Northern Caucasus, Karachai-Cherkessk Republic, N.A. Konstantinova & A.N. Savchenko, HRE 57 | MH367547 | * MH367781 | * MH367659 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | North America, Vancouver Island, B. Aguero, POZW 42444 | MH367549 | MH367783 | MH367661 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | USA, California Modoc Plateau, W.T. Doyle, MO 6005455 | MH367550 | * MH367784 | * MH367662 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | USA, Washington, W. Hong, MO 5241997 | MH367551 | * MH367785 | * MH367663 |
| Calypogeia azurea Stotler et Crotz | Calypogeia azurea Stotler et Crotz | USA, California, W.T. Doyle, 00234919 (NYBG) | MH367552 | * MH367786 | * MH367664 |
| Calypogeia cf. apiculata (Steph.) Steph. | Calypogeia cf. apiculata (Steph.) Steph. | Vietnam, Lao Cai Province, V.A. Bakalin, V-1-104-16 (VBGI) | OL689072 | - | - |
| Calypogeia cf. fissa (L.) Raddi | Calypogeia fissa (L.) Raddi | Georgia, Adjaria, V.A. Bakalin, G-12-70-13 (VBGI) | OL689083 | - | OL311474 |
| Calypogeia cf. integristipula Steph. | Calypogeia pseudointegristipula Bakalin, Troizk. et Maltseva | Russia, Sakhalin Province, Southern Kuril Islands, Kunashir Island, V.A. Bakalin & K.G. Klimova, K-34-34-18 (VBGI) | OL689071 | OL409006 | OL311448 |
| Calypogeia cf. japonica Steph. | Calypogeia sp. | Russia, Russian Far East, Southern Kurils, Kunashir Island, V.A. Bakalin & K.G. Klimova, K-35-22-18 (VBGI) | OL689074 | OL409008 | - |
| Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Poland, K. Buczkowska & A. Bączkiewicz, POZW 42211 | KR909089 | * KF371566 | * KF371618 |
| Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Poland, K. Buczkowska & A. Bączkiewicz, POZW 42220 | KR909087 | * KF371568 | * KF371620 |
| Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Poland, K. Buczkowska & A. Bączkiewicz, POZW 41707 | KR909086 | * KF371570 | * KF371622 |
| Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Poland, K. Buczkowska & A. Bączkiewicz, POZW 41706 | KR909085 | * KF371571 | * KF371623 |
| Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Poland, K. Buczkowska & A. Bączkiewicz, POZW 42285a | KR909088 | * KF371573 | * KF371625 |
| Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | Calypogeia cf. muelleriana (Schiffn.) Müll.Frib. | USA, B.K. Andreas, B.K.Andreas dhl825 | KR909090 | * KF371575 | * KF371627 |
| Calypogeia cf. sphagnicola f. paludosa (Warnst.) R.M. Schust. | Calypogeia cf. sphagnicola f. paludosa (Warnst.) R.M. Schust. | S Poland, POZW 41166 | - | * JQ658799 | JQ658782 |
| Calypogeia cf. tosana (Steph.) Steph. | Calypogeia japonica Steph. | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-1-31-17 (VBGI) | OL689036 | OL409013 | OL311467 |
| Calypogeia cuspidata (Steph.) Steph. | Calypogeia cuspidata (Steph.) Steph. | China, Sichuan Province, V.A. Bakalin & K.G. Klimova, C-44-1-17 (VBGI) | OL689041 | OL408967 | OL311478 |
| Calypogeia cuspidata (Steph.) Steph. | Calypogeia cuspidata (Steph.) Steph. | China, Sichuan Province, V.A. Bakalin & K.G. Klimova, C-40-6-17 (VBGI) | OL689038 | OL409015 | OL311480 |
| Calypogeia fissa (L.) Raddi | Calypogeia fissa (L.) Raddi | Germany, A. Solga, D. Quandt, POZW 39074 | - | MH367827 | * MH367701 |
| Calypogeia fissa (L.) Raddi | Calypogeia fissa (L.) Raddi | W Poland, Lubuskie Province, S. Rosadzinski, POZW 42437 | - | MH367828 | * MH367702 |
| Calypogeia fissa (L.) Raddi | Calypogeia fissa (L.) Raddi | Germany, A. Schäfer-Verwimp, S-V 25448 | - | MH367829 | MH367703 |
| Calypogeia fissa (L.) Raddi | Calypogeia fissa (L.) Raddi | North America, Canada, Nova Scotia, J. Macoun, POZW 3337 | - | MH367831 | MH367705 |
| Calypogeia fissa (L.) Raddi | Calypogeia aff. fissa (L.) Raddi | Ireland, J.S. Thomson, POZW 3344 | - | MH367832 | MH367706 |
| Calypogeia fissa (L.) Raddi | Calypogeia aff. fissa (L.) Raddi | Spain, C. Casas, POZW 3345 | - | MH367833 | MH367707 |
| Calypogeia fissa (L.) Raddi | Calypogeia fissa (L.) Raddi | Hungary, L. Vajda, POZW 3347 | - | MH367834 | MH367708 |
| Calypogeia fissa (L.) Raddi | Calypogeia aff. fissa (L.) Raddi | Iceland, Austurnssysla, V.V. Buryy, Ice-3-3-17 | - | OL408960 | OL311418 |
| Calypogeia fissa (L.) Raddi | Calypogeia shevockii Bakalin et Troizk. | USA, California State, J.R. Shevock, 50287 | OL689039 | OL408965 | OL311477 |
| Calypogeia fissa (L.) Raddi | Calypogeia fissa (L.) Raddi | Georgia, Adjaria, V.A. Bakalin, G-12-55-13 (VBGI) | OL689037 | OL408968 | OL311473 |
| Calypogeia fissa (L.) Raddi | Asperifolia sullivantii (Austin) Troizk., Bakalin, Maltseva | USA, Sainte Genevieve County, V.A. Bakalin, US-41a-80-14 (VBGI) | OL689045 | OL408971 | OL409018 |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, S. Rosadziński, POZW 42628 | KR909073 | * KR909061 | - |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, K. Buczkowska & J. Szweykowski, POZW 39203 | KR909074 | * KR909062 | - |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, K. Buczkowska, POZW 42225 | KR909075 | * KR909063 | - |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, K. Buczkowska, POZW 42227 | KR909076 | * KR909064 | - |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, K. Buczkowska & A. Bączkiewicz, POZW 42345 | KR909077 | * KR909065 | - |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, K. Buczkowska & A. Bączkiewicz, POZW 42205 | KR909078 | * KR909066 | - |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, K. Buczkowska & A. Bączkiewicz, POZW 42200 | KR909079 | * KR909067 | - |
| Calypogeia fissa (L.) Raddi subsp. fissa | Calypogeia fissa (L.) Raddi subsp. fissa | Poland, S. Rosadziński & K. Buczkowska, POZW 42298 | KR909080 | * KR909068 | - |
| Calypogeia fissa subsp. neogaea R.M. Schust. | Calypogeia neogaea (R.M. Schust.) Bakalin | USA, North Carolina, B. Shaw, POZW 42620 | KR909081 | KR909069 | - |
| Calypogeia fissa subsp. neogaea R.M. Schust. | Calypogeia neogaea (R.M. Schust.) Ba-kalin | USA, North Carolina, B. Shaw, POZW 42622 | KR909082 | KR909070 | - |
| Calypogeia fissa subsp. neogaea R.M. Schust. | Calypogeia neogaea (R.M. Schust.) Bakalin | USA, North Carolina, B. Shaw, POZW 42625 | KR909083 | KR909071 | - |
| Calypogeia fissa subsp. neogaea R.M. Schust. | Calypogeia neogaea (R.M. Schust.) Bakalin | USA, North Carolina, B. Shaw, POZW 42626 | KR909084 | KR909072 | - |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | Vietnam, Lai Châu Province, V.A. Bakalin & K.G. Klimova, V-13-8-19 (VBGI) | OL689076 | - | OL311419 |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | Vietnam, Sơn La Province, V.A. Bakalin & K.G. Klimova, V-21-16-19 (VBGI) | OL689080 | - | OL311420 |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | Vietnam, Lai Châu Province, V.A. Bakalin & K.G. Klimova, V-3-4-19 (VBGI) | OL689084 | - | OL311421 |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | Vietnam, Lai Châu Province, V.A. Bakalin & K.G. Klimova, V-8-4-19 (VBGI) | OL689034 | - | OL311422 |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | Japan, H. Inoue, JE 18004 | MH367564 | MH367802 | MH367680 |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | China, Guizhou Province, V.A. Bakalin, China-56-19-13 (VBGI) | MH367566 | MH367804 | * MH367682 |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | Vietnam, Hà Giang Province, V.A. Bakalin & K.G. Klimova, V-11-24-20 (VBGI) | OL689050 | OL770363 | OL770364 |
| Calypogeia granulata Inoue | Calypogeia granulata Inoue | Japan, Kuroyama, H. Inoue, 2792205 (NYGB) | MH367565 | * MH367803 | * MH367681 |
| Calypogeia integristipula Steph. | Calypogeia integristipula Steph. | Finland, He-Nygrèn & Piippo 1472 | - | AY463550 | - |
| Calypogeia integristipula Steph. | Calypogeia integristipula Steph. | NE Poland, K. Buczkowska & A. Bączkiewicz, POZW 41730 | - | MH367823 | * MH367697 |
| Calypogeia integristipula Steph. | Calypogeia integristipula Steph. | SE Poland, K. Buczkowska & B. Chmielewski, POZW 41928 | - | MH367824 | * MH367698 |
| Calypogeia integristipula Steph. | Calypogeia integristipula Steph. | NE Poland, K. Buczkowska & A. Bączkiewicz, POZW 41785 | - | MH367825 | * MH367699 |
| Calypogeia integristipula Steph. | Calypogeia integristipula Steph. | NW Poland, K. Buczkowska & A. Bączkiewicz, POZW 41188 | - | MH367826 | MH367700 |
| Calypogeia integristipula Steph. | Calypogeia integristipula Steph. | Russia, Russian Far East, Khabarovsk Territory, V.A. Bakalin, Kh-28-7-16 (VBGI) | OL689044 | OL408970 | OL311423 |
| Calypogeia japonica Steph. | Calypogeia japonica Steph. | Japan, Honshu Island, V.A. Bakalin, J-1-15-13 (VBGI) | - | OL408974 | OL311424 |
| Calypogeia japonica Steph. | Calypogeia japonica Steph. | Japan, Shikoku Island, V.A. Bakalin, J-4-3-15 (VBGI) | OL689047 | OL408975 | OL311476 |
| Calypogeia japonica Steph. | Calypogeia japonica Steph. | Japan, Kyushu Island, V.A. Bakalin, J-7-30-14 (VBGI) | OL689048 | OL408976 | OL311425 |
| Calypogeia japonica Steph. | Calypogeia sp. | Vietnam, Lao Cai Province, V.A. Bakalin, V-4-22-16 (VBGI) | OL689053 | OL408981 | - |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia muelleriana (Schiffn.) Müll.Frib. | Russia, Perm Province, N. Konstantinova, K 367-1-04 (KPABG) | JX629935 | * JX630062 | - |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia muelleriana (Schiffn.) Müll.Frib. | Poland, K. Buczkowska & A. Bączkiewicz, POZW 41346 | - | KF371550 | * KF371602 |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia muelleriana (Schiffn.) Müll.Frib. | Poland, K. Buczkowska, A. Baczkiewicz, POZW 41182 | - | * KF371551 | KF371603 |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia muelleriana (Schiffn.) Müll.Frib. | Netherlands, J. Szweykowski, R. Gradstein, H. Greven, POZW 34160 | - | KF371563 | * KF371615 |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia muelleriana (Schiffn.) Müll.Frib. | Russia, Russian Far East, Kamchatka Territory, V.A. Bakalin, K-50-2-15 (VBGI) | - | OL408982 | - |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia pseudosphagnicola Bakalin, Troizk. et Maltseva | Russia, Russian Far East, Khabarovsk Territory, V.A. Bakalin, Kh-23-2-16 (VBGI) | OL689054 | OL408983 | OL311453 |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia muelleriana (Schiffn.) Müll.Frib. | Russia, Russian Far East, Magadan Province, V.A. Bakalin, Mag-31-26-12 (VBGI) | - | OL408984 | - |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | Russia, Russian Far East, Magadan Province, V.A. Bakalin, Mag-32-41a-12 (VBGI) | - | OL408985 | OL311455 |
| Calypogeia muelleriana (Schiffn.) Müll.Frib. | Calypogeia pseudointegristipula Bakalin, Troizk. et Maltseva | Russia, Russian Far East, Sakhalin Province, Sakhalin Island, V.A. Bakalin, S-43-6-16 (VBGI) | - | OL408986 | OL311457 |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Poland, POZW 41952 | - | MK294001 | - |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Poland, K. Buczkowska, A. Baczkiewicz, POZW 41735 | - | MH367820 | * MH367694 |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Poland, K. Buczkowska, B. Chmielewski, POZW 41927 | - | MH367821 | * MH367695 |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Poland, K. Buczkowska, A. Baczkiewicz, POZW 41358 | - | MH367822 | * MH367696 |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Poland, POZW 41731 | - | MK294002 | - |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Poland, K. Buczkowska, A. Baczkiewicz, POZW 41731 | - | MH367819 | MH367693 |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Russia, Russian Far East, Sakhalin Province, Sakhalin Island, S.V. Dudov, 2016_Br_0073 | - | OL408987 | OL311434 |
| Calypogeia neesiana (C.Massal. et Carestia) Müll.Frib. | Calypogeia muelleriana (Schiffn.) Müll. Frib. | Russia, Russian Far East, Sakhalin Province, Sakhalin Island, S.V. Dudov & K.V. Kotelnikova, 2014_Br_0205 | - | OL408988 | OL409017 |
| Calypogeia neesiana ssp. subalpina (Inoue) Inoue | Calypogeia subalpina Inoue | Japan, Honshu Island, V.A. Bakalin, J-87-1-15 (VBGI) | - | OL408993 | OL311450 |
| Calypogeia neogaea (R.M. Schust.) Bakalin | Calypogeia kamchatica Bakalin, Troizk. et Maltseva | Russia, Russian Far East, Kamchatka Territory, V.A. Bakalin, HRE, 28 (VBGI) | OL689060 | OL408994 | OL311445 |
| Calypogeia neogaea (R.M. Schust.) Bakalin | Calypogeia kamchatica Bakalin, Troizk. et Maltseva | Russia, Russian Far East, Kamchatka Territory, V.A. Bakalin, K-63-1-15 (VBGI) | OL689061 | OL408995 | OL311446 |
| Calypogeia neogaea (R.M. Schust.) Bakalin | Calypogeia tosana (Steph.) Steph. | Russia, Russian Far East, Sakhalin Province, Iturup Island, V.A. Bakalin, K-72-13-15 (VBGI) | OL689062 | OL408996 | OL311452 |
| Calypogeia neogaea (R.M. Schust.) Bakalin | Calypogeia neogaea (R.M. Schust.) Bakalin | USA, Sainte Genevieve County, V.A. Bakalin, US-41-77-14 (VBGI) | OL689063 | OL408997 | OL311428 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Russia, Russian Far East, Primorsky Territory, V.A. Bakalin, P-40-1-12 (VBGI) | - | * MH367790 | MH367668 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Japan, M. Higuchi, 02532867 (NYBG) | - | * MH367795 | MH367673 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Japan, K. Yamada, Yamada 8912 | - | * MH367796 | MH367674 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Russia, Russian Far East, Primorsky Territory, V.A. Bakalin, P-15-12-12 (VBGI) | MH367553 | MH367788 | MH367666 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Russia, Russian Far East, Primorsy Territory, V.A. Bakalin, P-39-4-12 (VBGI) | MH367554 | MH367789 | MH367667 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Korea, Gangwon Province, V.A. Bakalin, Kor-7-23-11 (VBGI) | MH367555 | * MH367791 | * MH367669 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Korea, Gangwon Province, V.A. Bakalin, Kor-10-02-11 (VBGI) | MH367556 | * MH367792 | MH367670 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Korea, Gangwon Province, V.A. Bakalin, Kor-7-36-11 (VBGI) | MH367557 | * MH367793 | MH367671 |
| Calypogeia orientalis Buczkowska et Bakalin | Calypogeia orientalis Buczkowska et Bakalin | Japan, Kyushu Island, V.A. Bakalin, J-7-79-14 (VBGI) | MH367558 | * MH367794 | MH367672 |
| Calypogeia peruviana Nees et Mont. | Calypogeia peruviana Nees et Mont. | USA, North Carolina, B. Aguero, POZW 42619 | MH367562 | MH367800 | MH367678 |
| Calypogeia peruviana Nees et Mont. | Calypogeia peruviana Nees et Mont. | USA, North Carolina, B. Aguero, POZW 42627 | MH367563 | MH367801 | * MH367679 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | Vietnam, Sơn La Province, V.A. Bakalin & K.G. Klimova, V-21-26-19 (VBGI) | - | - | OL311429 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | Vietnam, Sơn La Province, V.A. Bakalin & K.G. Klimova, V-23-15-19 (VBGI) | - | - | OL311430 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | China, Guizhou Proivince, V.A. Bakalin, China-56-77-13 (VBGI) | MH367559 | MH367797 | MH367675 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | China, Guizhou Proivince, V.A. Bakalin, China-56-78-13 (VBGI) | MH367560 | * MH367798 | MH367676 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | Vietnam, Lao Cai Province, V.A. Bakalin, V-2-73-16 (VBGI) | MH367561 | * MH367799 | MH367677 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | Vietnam, Hà Giang Province, V.A. Bakalin & K.G. Klimova, V-11-7-20 (VBGI) | OL689032 | OL408962 | OL311462 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | Vietnam, Cao Bằng Province, V.A. Bakalin & K.G. Klimova, V-27-4-20 (VBGI) | OL689035 | OL408964 | OL311469 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | Vietnam, Cao Bằng Province, V.A. Bakalin & K.G. Klimova, V-24-10-20 (VBGI) | OL689078 | OL409011 | OL311468 |
| Calypogeia sinensis Bakalin et Buczkowska | Calypogeia sinensis Bakalin et Buczkowska | Vietnam, Hà Giang Province, V.A. Bakalin & K.G. Klimova, V-11-23-20 (VBGI) | OL689082 | OL409014 | OL311461 |
| Calypogeia sp. nov. | Calypogeia granulata Inoue | Vietnam, Lao Cai Province, V.A. Bakalin, V-1-130-16 (VBGI) | OL689042 | - | OL311447 |
| Calypogeia sp. | Calypogeia apiculata (Steph.) Steph. | Vietnam, Lai Châu Province, V.A. Bakalin & K.G. Klimova, V-12-5-19 (VBGI) | OL689043 | OL408969 | OL311464 |
| Calypogeia sp. | Calypogeia pseudocuspidata Bakalin, Frank Müll. et Troizk. | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-21-17-18 (VBGI) | OL689046 | OL408972 | OL311458 |
| Calypogeia sp. | Calypogeia sp. | Russia, Russian Far East, Primorsky Territory, V.A. Bakalin, P-37-20-14 | - | OL408973 | - |
| Calypogeia sp. | Calypogeia muelleriana (Schiffn.) Müll.Frib. | Russia, Russian Far East, Khabarovsk Territory, K.G. Klimova, Khab-69-31-18 (VBGI) | OL689049 | OL408977 | - |
| Calypogeia sp. | Calypogeia apiculata (Steph.) Steph. | Vietnam, Lai Châu Province, V.A. Bakalin & K.G. Klimova, V-4-15-19 (VBGI) | OL689051 | OL408979 | OL311459 |
| Calypogeia sp. | Calypogeia sp. | Japan, Honshu Island, M. Higuchi, BSE, 1293 | OL689059 | OL408992 | - |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Calypogeia pseudosphagnicola Bakalin, Troizk. et Maltseva | Russia, Russian Far East, Khabarovsk Territory, V.A. Bakalin, Kh-40-28-13 (VBGI) | - | - | OL311454 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Russia, Russian Far East, Khabarovsk Territory, K.G. Klimova, Khab-68-14-18 (VBGI) | - | - | OL311432 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Russia, Russian Far East, Magadan Province, V.A. Bakalin, Mag-38-16-12 (VBGI) | - | - | OL311433 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Calypogeia sp. | Japan, H. Inoue, KRAM 50656 | - | - | MH367709 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Russia, Russian Far East, Kamchatka Territory, V.A. Bakalin, K-54-1-15 (VBGI) | - | OL408998 | OL311431 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske | Calypogeia pseudosphagnicola Bakalin, Troizk. et Maltseva | Russia, Russian Far East, Sakhalin Province, Sakhalin Island, V.A. Bakalin, S-17-3-17 (VBGI) | - | OL408999 | OL311456 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | NW Poland, K. Buczkowska & A. Bączkiewicz, POZW 42284 | - | JQ658790 | JQ658773 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | NW Poland, K. Buczkowska & A. Bączkiewicz, POZW 42245 | - | JQ658791 | * JQ658774 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | Poland, K. Buczkowska & A. Bączkiewicz, POZW 42351 | - | JQ658792 | * JQ658775 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | NW Poland, K. Buczkowska & A. Bączkiewicz, POZW 42266 | - | JQ658793 | * JQ658776 |
| Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | Calypogeia sphagnicola (Arnell et J.Perss.) Warnst. et Loeske f. sphagnicola | NE Poland, K. Buczkowska & A. Bączkiewicz, POZW 41711 | - | JQ658794 | JQ658777 |
| Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | S Poland, K. Buczkowska & A. Bączkiewicz, POZW 41174 | - | JQ658795 | JQ658778 |
| Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | S Poland, K. Buczkowska & A. Bączkiewicz, POZW 41148 | - | JQ658796 | JQ658779 |
| Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | S Poland, K. Buczkowska & A. Bączkiewicz, POZW 42277 | - | JQ658797 | * JQ658780 |
| Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | Calypogeia sphagnicola f. paludosa (Warnst.) Schust. | S Poland, K. Buczkowska & A. Bączkiewicz, POZW 41722 | - | JQ658798 | * JQ658781 |
| Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Poland, J. Szweykowski, K. Buczkowska, POZW 39500 | - | * MH367815 | MH367689 |
| Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Russia, Russian Far East, Khabarovsk Territory, K.G. Klimova, Khab-60-34-18 (VBGI) | - | - | OL311436 |
| Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Calypogeia sp. | Russia, Russian Far East, Primorsky Territory, V.A. Bakalin & G. Arutinov, P-2-6a-13 (VBGI) | - | - | OL311460 |
| Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Calypogeia suecica (Arnell et J.Perss.) Müll. Frib. | Russia, Russian Far East, Primorsky Territory, V.A. Bakalin, P-37-1-14 (VBGI) | - | - | OL311437 |
| Calypogeia sullivantii Austin | Asperifolia sullivantii (Austin) Troizk., Bakalin, Maltseva | USA, North Carolina, B. Aguero, POZW 42623 | MH367569 | MH367813 | MH367687 |
| Calypogeia sullivantii Austin | Asperifolia sullivantii (Austin) Troizk., Bakalin, Maltseva | USA, Maryland, L.T. Biechele, NYGB 2169286 | MH367570 | MH367814 | * MH367688 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | China, Guizhou Province, V.A. Bakalin, C-55-72-13 (VBGI) | OL689067 | - | OL311438 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | China, Guizhou Proivince, V.A. Bakalin, China-56-86-13 (VBGI) | - | MH367807 | * MH367683 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Japan, Kyushu Island, Fukuoka Prefecture, V.A. Bakalin, J-4-38-14 (VBGI) | - | MH367808 | MH367684 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Vietnam, Lao Cai Province, V.A. Bakalin, V-1-102-16 (VBGI) | OL689033 | OL408963 | - |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Vietnam, Cao Bằng Province, V.A. Bakalin & K.G. Klimova, V-23-2-20 (VBGI) | OL689040 | OL408966 | OL311471 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | China, Guizhou Province, V. A. Bakalin, C-55-19-13 (VBGI) | - | OL408978 | OL311479 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Vietnam, Lai Châu Province, V.A. Bakalin & K.G. Klimova, V-16-2-19 (VBGI) | OL689052 | OL408980 | OL311440 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Republic of Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-73-16-19 (VBGI) | OL689058 | OL408991 | OL311435 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Republic of Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-79-25-19 (VBGI) | OL689066 | OL409002 | OL311439 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia yoshinagana Steph. | Japan, Kyushu Island, V.A. Bakalin, J-40-52-14 (VBGI) | OL689068 | OL409003 | OL311449 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Russia, Russian Far East, Sakhalin Province, Iturup Island, V.A. Bakalin, K-70-35-15 (VBGI) | OL689069 | OL409004 | OL311451 |
| Calypogeia tosana (Steph.) Steph. | Calypogeia yoshinagana Steph. | Republic Of Korea, Jeju Island, V.A. Bakalin, Kor-29-64-15 (VBGI) | OL689070 | OL409005 | - |
| Calypogeia tosana (Steph.) Steph. | Calypogeia tosana (Steph.) Steph. | Japan, Honshu Island, V.A. Bakalin, J-5-9-13 (VBGI) | - | OL409009 | - |
| Calypogeia vietnamica Bakalin et Vilnet | Calypogeia vietnamica Bakalin et Vilnet | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-8-61-17 (VBGI) | MK335824 | MK336253 | - |
| Calypogeia vietnamica Bakalin et Vilnet | Calypogeia vietnamica Bakalin et Vilnet | Vietnam, Lao Cai Province, V.A. Bakalin & K.G. Klimova, V-9-23-17 (VBGI) | MK335825 | MK336254 | - |
| Calypogeia yoshinagana Steph. | Calypogeia yoshinagana Steph. | Republic of Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-72-5-19 (VBGI) | OL689075 | - | OL311442 |
| Calypogeia yoshinagana Steph. | Calypogeia yoshinagana Steph. | Republic of Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-79-22-19 (VBGI) | OL689077 | - | OL311443 |
| Calypogeia yoshinagana Steph. | Calypogeia yoshinagana Steph. | Republic of Korea, Jeollabuk-do, V.A. Bakalin & S.S. Choi, Kor-82-3-19 (VBGI) | OL689081 | - | OL311444 |
| Calypogeia yoshinagana Steph. | Calypogeia yoshinagana Steph. | Republic of Korea, Gyeongsangnam-do, V.A. Bakalin & S.S. Choi, Kor-70-1-19 (VBGI) | OL689073 | OL409007 | OL311441 |
| Geocalyx graveolens (Schrad.) Nees | Geocalyx graveolens (Schrad.) Nees | China, Yunnan Province, D.G. Long, 34828 (E) | - | * KJ802067 | KJ802038 |
| Mesoptychia sahlbergii (Lindb. et Arnell) A. Evans | Mesoptychia sahlbergii (Lindb. et Arnell) A. Evans | USA, D.G. Long, 11329 (E) | KJ206417 | - | - |
| Mesoptychia gillmanii (Austin) L. Söderstr. et Váňa | Mesoptychia gillmanii (Austin) L. Söderstr. et Váňa | Sweden, A. Cailliau & N. Lonnell, 982 (G) | KJ206404 | * KJ206363 | - |
| Mesoptychia gillmanii (Austin) L. Söderstr. et Váňa | Mesoptychia gillmanii (Austin) L. Söderstr. et Váňa | Russia, N.A. Konstantinova & A.N. Savchenko, 117-1-00 (F) | - | * KJ802075 | KJ802047 |
| Mesoptychia heterocolpos (Thed. ex Hartm.) L. Söderstr. et Váňa | Mesoptychia heterocolpos (Thed. ex Hartm.) L. Söderstr. et Váňa | USA, Alaska, B. Shaw, F965a/3 (DUKE) | - | * KF943057 | KF942896 |
| Mesoptychia heterocolpos (Thed. ex Hartm.) L. Söderstr. et Váňa | Mesoptychia heterocolpos (Thed. ex Hartm.) L. Söderstr. et Váňa | Sweden, A. Cailliau, 999 (G) | KJ206411 | * KJ206371 | - |
| Mesoptychia heterocolpos (Thed. ex Hartm.) L. Söderstr. et Váňa | Mesoptychia heterocolpos (Thed. ex Hartm.) L. Söderstr. et Váňa | Russia, Komi Rep., Dulin M.V., 116749 (KPABG) | - | * KM501496 | KM501477 |
| Mesoptychia heterocolpos var. harpanthoides (Bryhn et Kaal.) L. Söderstr. et Váňa | Mesoptychia heterocolpos var. harpanthoides (Bryhn et Kaal.) L. Söderstr. et Váňa | Russia, Arkhangelsk Prov., Franz Josef Land, Ziegler Island, A. Savchenko, CA-19-29-8c (KPABG) | - | * MT431408 | MT431398 |
| Mesoptychia sahlbergii (Lindb. et Arnell) A. Evans | Mesoptychia sahlbergii (Lindb. et Arnell) A. Evans | Russia, V. Fedosov, 107967 (F) | - | * KJ802078 | KJ802050 |
| Metacalypogeia alternifolia (Nees) Grolle | Metacalypogeia alternifolia (Nees) Grolle | Bhutan, D.G. Long, 28712 (E) | - | KJ802068 | KJ802040 |
| Metacalypogeia cordifolia (Steph.) Inoue | Metacalypogeia cordifolia (Steph.) Inoue | Russia, Russian Far East, Primorsky Territory, V.A. Bakalin, P-66-18a-06 (VBGI) | * JX629934 | JF421597 | - |
| Mnioloma fuscum (Lehm.) R.M. Schust. | Mnioloma fuscum (Lehm.) R.M. Schust. | South Africa, de Roo, s.n. (BOL) | - | - | AM397718 |
| Mnioloma fuscum (Lehm.) R.M. Schust. | Mnioloma fuscum (Lehm.) R.M. Schust. | Fiji, J.E. Braggins et al. 16 Apr 2008 (NSW) | - | KJ802100 | - |
| Taxon | Intergeneric p-Distances, trnG-Intron/trnL−F/ITS2 | ||||
|---|---|---|---|---|---|
| Calypogeia | Asperifolia | Mnioloma | Metacalypogeia | Geocalyx | |
| Asperifolia | 0.17/0.13/0.13 | ||||
| Mnioloma | 0.17/0.16/- | 0.17/0.16/- | |||
| Metacalypogeia | 0.15/0.12/- | 0.21/0.15/- | 0.20/0.16/- | ||
| Geocalyx | 0.15/0.17/- | 0.2/0.18/- | 0.19/0.18/- | 0.18/0.14/- | |
| Mesoptychia | 0.16/0.15/0.19 | 0.19/0.17/0.22 | 0.18/0.19/- | 0.19/0.13/- | 0.16/0.13/- |
| Locus | Sequence (5′-3′) | Direction | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| ITS2 nrDNA | GCATCGATGAAGAACGCAGC | forward | 62 | [33] |
| ITS2 nrDNA | GATATGCTTAAACTCAGCGG | reverse | 58 | [33] |
| ITS2 nrDNA | GAGTCATCAGCTCGCGTTGAC | forward | 66 | [34] |
| ITS2 nrDNA | GCTGCGTTCTTCATCGATGC | reverse | 62 | [35] |
| trnL–F cpDNA | CGAATTCGGTAGACGCTACG | forward | 62 | [36] |
| trnL–F cpDNA | ATTTGAACTGGTGACACGAG | reverse | 58 | [36] |
| trnL–F cpDNA | CGAAATTGGTAGACGCTGCG | forward | 62 | [37] |
| trnL–F cpDNA | TGCCAGAAACCAGATTTGAAC | reverse | 60 | [37] |
| trnG-intron cpDNA | ACCCGCATCGTTAGCTTG | forward | 56 | [38] |
| trnG-intron cpDNA | GCGGGTATAGTTTAGTGG | reverse | 54 | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalin, V.A.; Maltseva, Y.D.; Müller, F.; Klimova, K.G.; Nguyen, V.S.; Choi, S.S.; Troitsky, A.V. Calypogeia (Calypogeiaceae, Marchantiophyta) in Pacific Asia: Updates from Molecular Revision with Particular Attention to the Genus in North Indochina. Plants 2022, 11, 983. https://doi.org/10.3390/plants11070983
Bakalin VA, Maltseva YD, Müller F, Klimova KG, Nguyen VS, Choi SS, Troitsky AV. Calypogeia (Calypogeiaceae, Marchantiophyta) in Pacific Asia: Updates from Molecular Revision with Particular Attention to the Genus in North Indochina. Plants. 2022; 11(7):983. https://doi.org/10.3390/plants11070983
Chicago/Turabian StyleBakalin, Vadim A., Yulia D. Maltseva, Frank Müller, Ksenia G. Klimova, Van Sinh Nguyen, Seung Se Choi, and Aleksey V. Troitsky. 2022. "Calypogeia (Calypogeiaceae, Marchantiophyta) in Pacific Asia: Updates from Molecular Revision with Particular Attention to the Genus in North Indochina" Plants 11, no. 7: 983. https://doi.org/10.3390/plants11070983