

Hidden Diversity within Tetralophozia filiformis (Marchantiophyta, Anastrophyllaceae) in East Asia
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Molecular Genetic
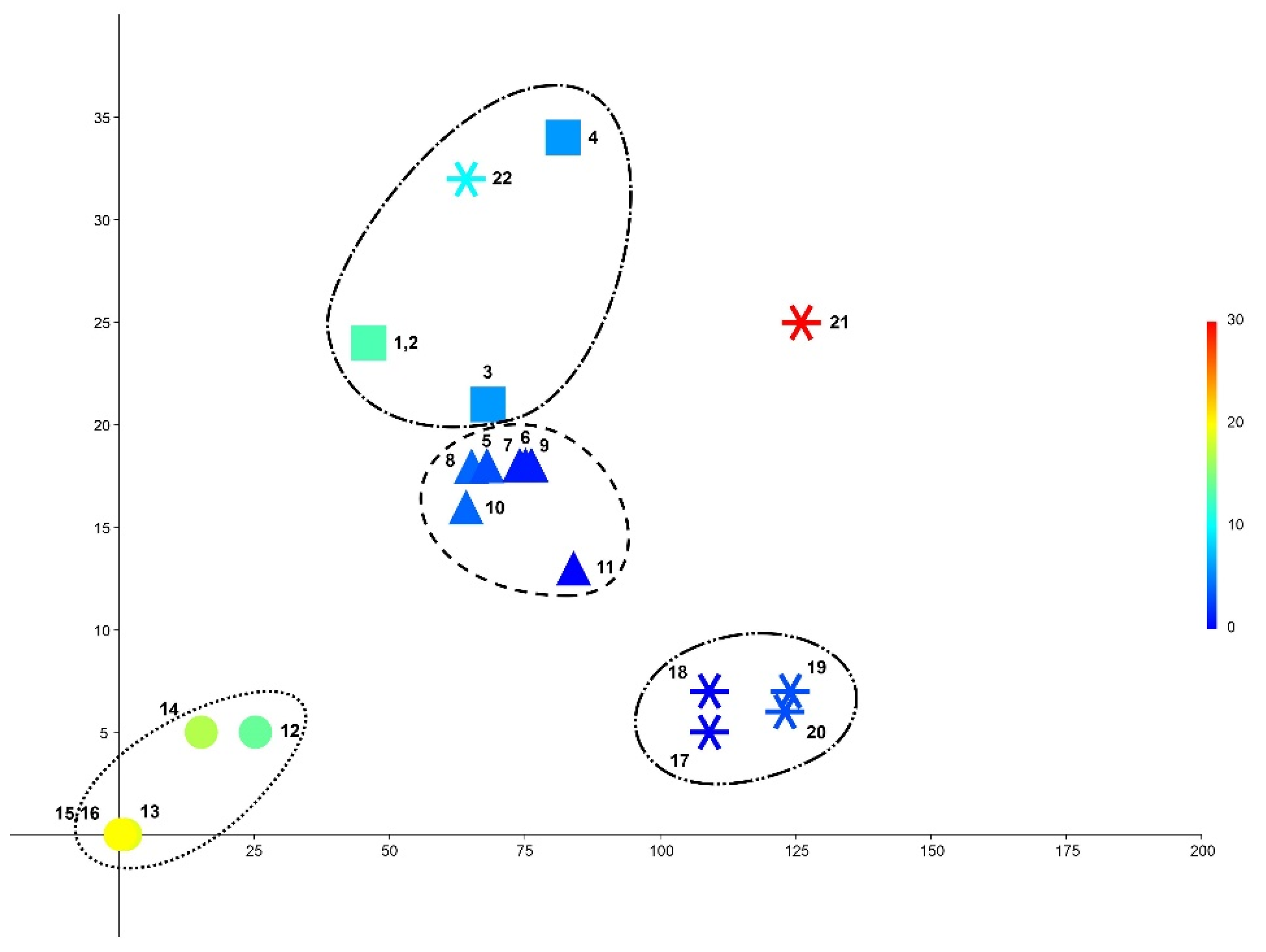
2.2. Climate Variables and the Genetic Diversification
- Tetralophozia pusilla and T. filiformis are closely related in the molecular-genetic respect and grow in similar climatic environments too. Tetralophozia sibirica grows in harsh northern environments and is genetically well different from other two taxa that also correspond distance in climate diagram.
- Strongly geographically distanced Alaskan and Spanish populations were found to be similar in measured climate variables.
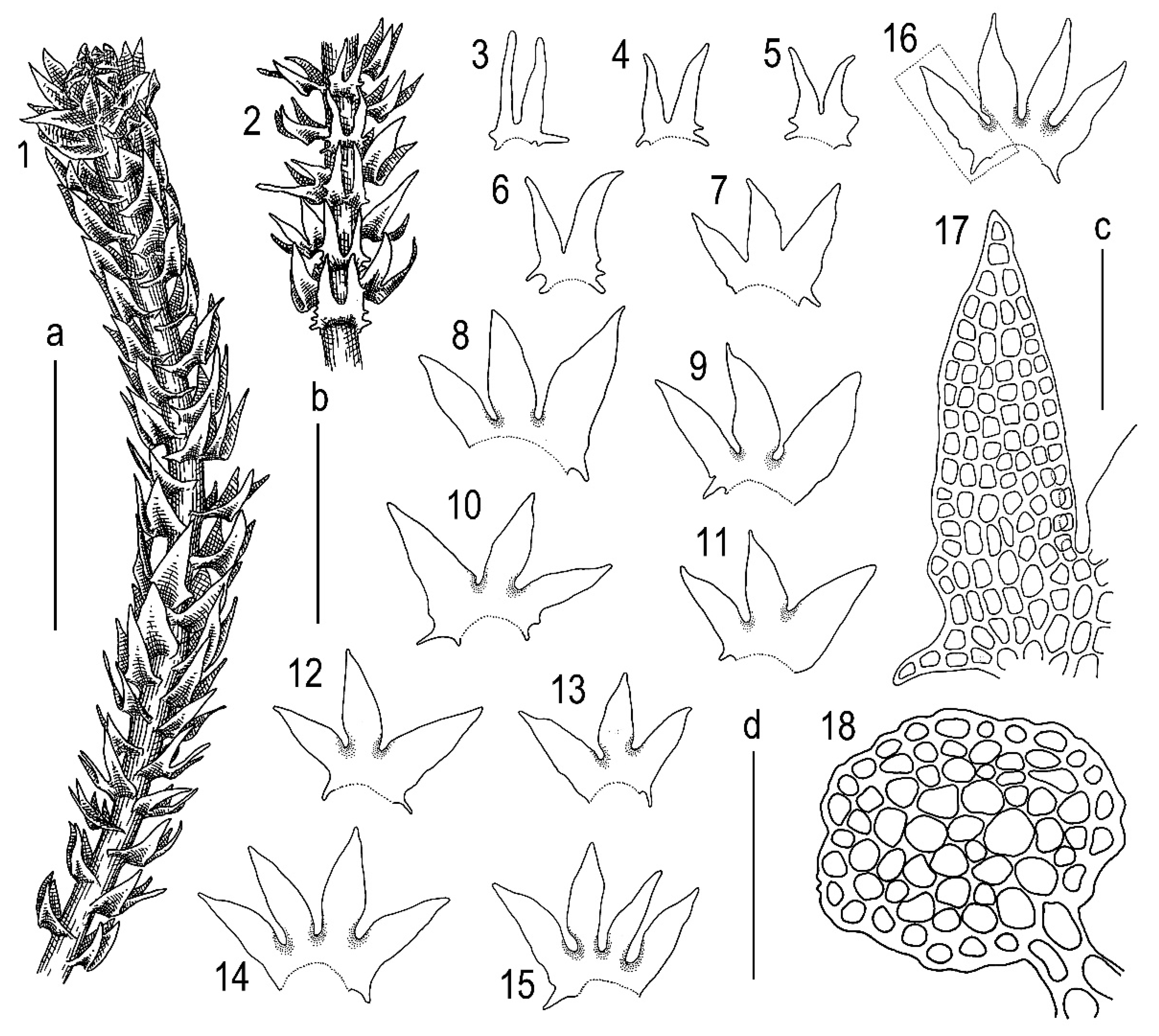
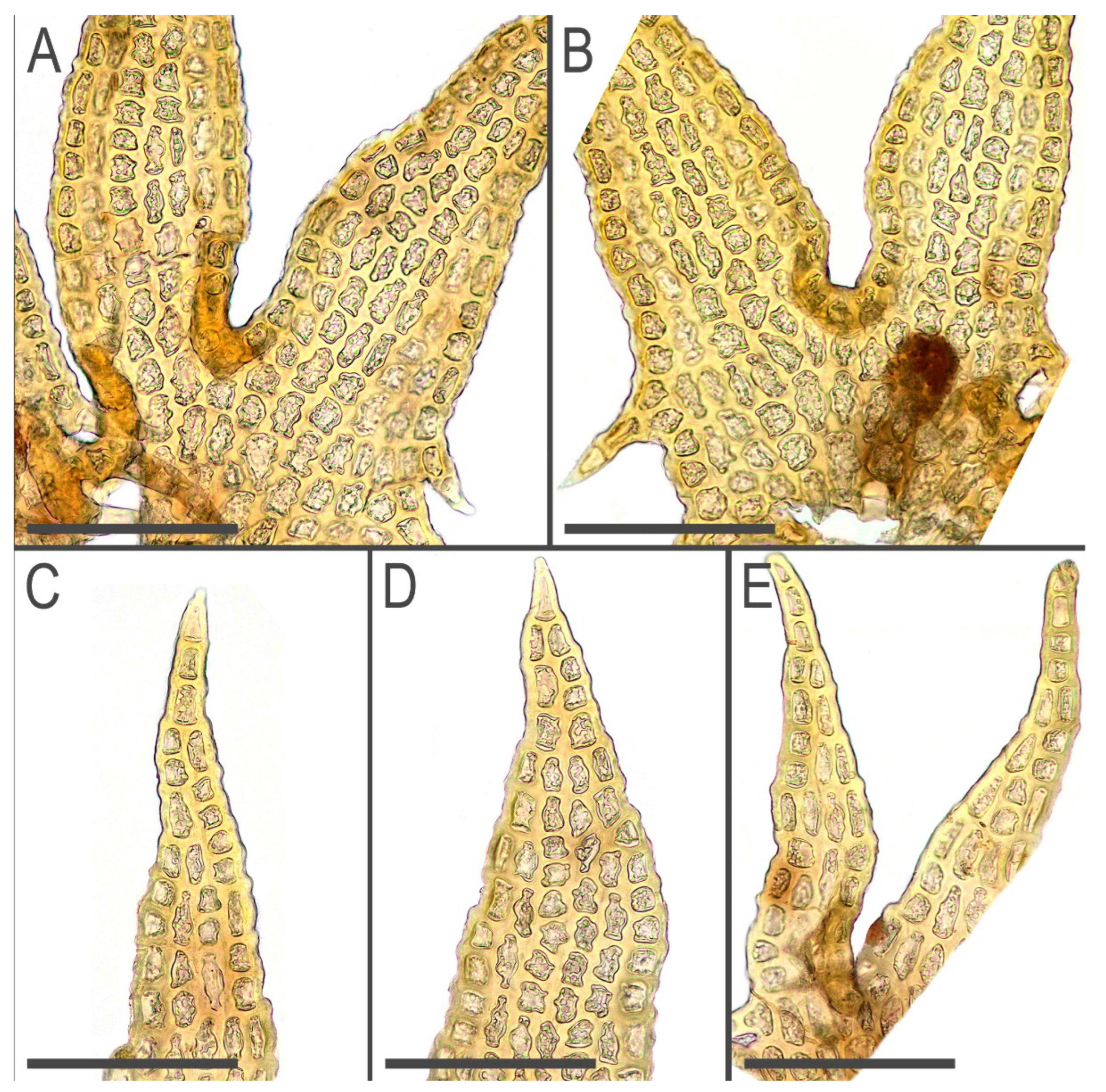
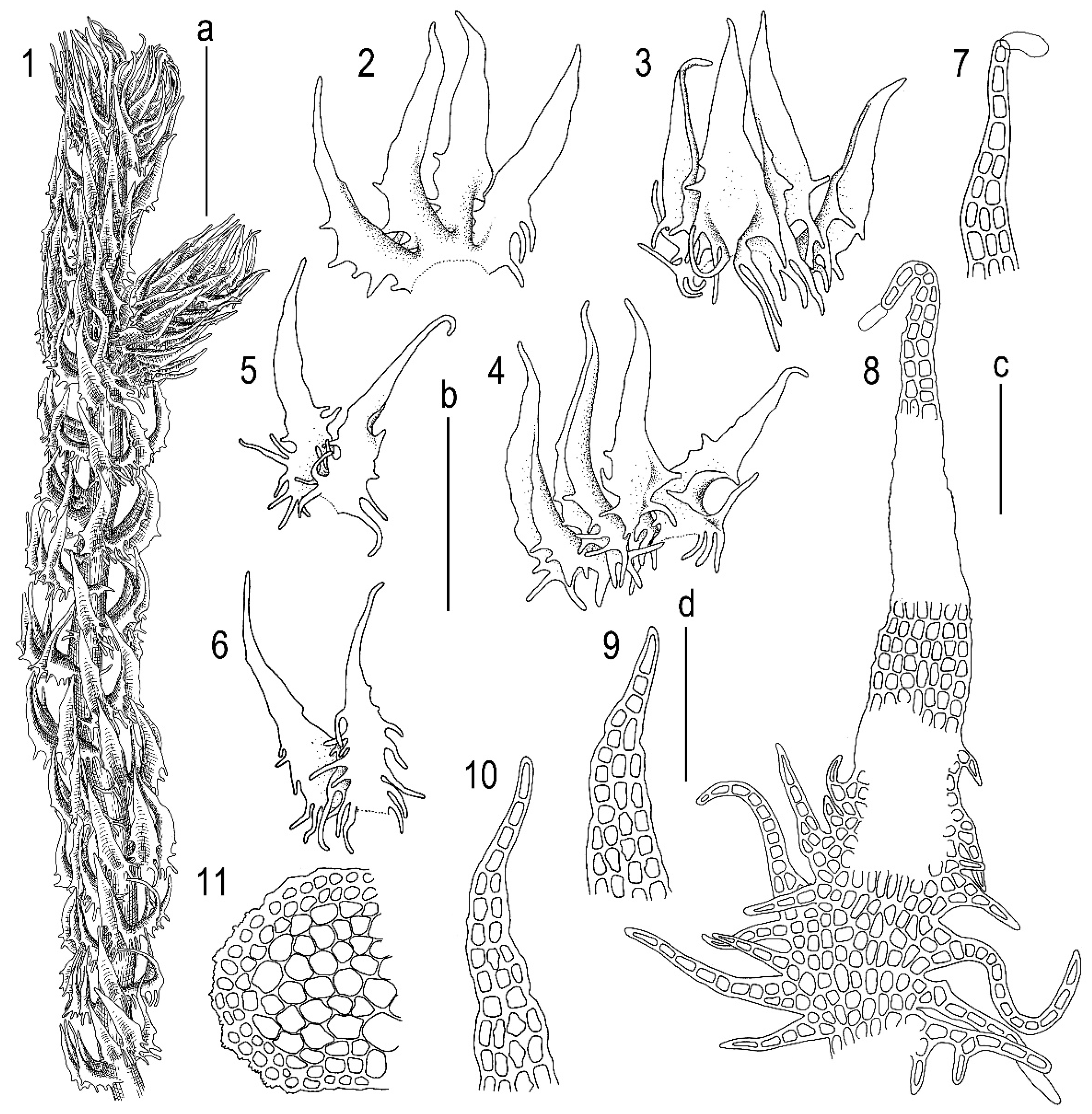
2.3. Taxonomy
3. Discussion
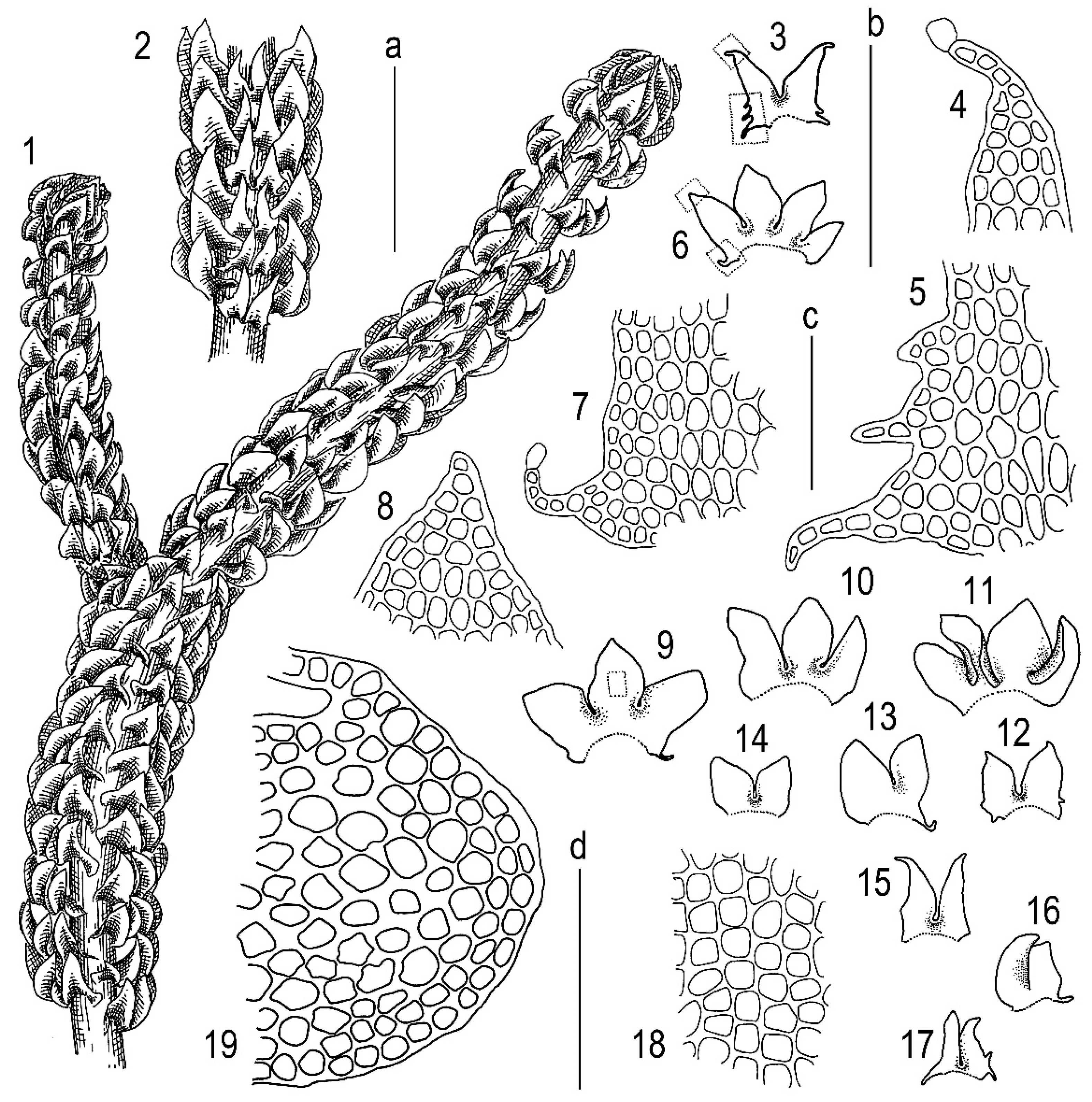
3.1. Morphology
3.2. The Distribution of ‘Narrow’ Taxa of Tetralophozia filiformis Complex
Key to Tetralophozia in Amphi-Pacific Asia
- 1.
- Plants commonly more than 0.7–0.8 mm wide (depauperate modifications with commonly bilobed leaves, including ‘f. alpina’ are narrower), leaf middle lobe 1.5–2.2 as long as wide, leaf cuticle virtually smooth [arctic-alpine circumpolar] … T. setiformis *
- 1.
- Plants less than 0.7–0.8 mm wide, leaf middle lobe (2.5–)3–4 as long as wide, terminating by 1-several uniseriate cells, leaf cuticle papillose-verrucose, sometimes scarcely or coarsely so … 2.
- 2.
- Plants 0.3–0.5 mm wide, cells in the lobe base 12–30 × 12–15 µm, cuticle papillose, sometimes obscurely so, leaf basal teeth less 10 cells long [hemiboreal Asian mainland with continental climate] … T. sibirica
- 2.
- Plants wider, 0.6–0.75 mm wide, or almost the same size (0.4–0.6 mm wide), but then with cells in the lobe base are distinctly smaller (10–20 × 10–12 µm), leaf basal teeth less than 10 cells long or to 20 cells long [warm-temperate to tropical East Asian mainland or temperate insular-peninsular East Asia, monsoon to oceanic climates] … 3
- 3.
- Plants 0.6–0.75 mm wide, leaf undivided part 30–50 µm (2–5 cells) high, cells in the leaf lobe base 12–25 × (7–)10–12(–15) µm, cuticle distinctly papillose-verrucose, sometimes coarsely so [warm-temperate to tropical East and SE Asia] … T. filiformis
- 3.
- Plants 0.4–0.6 mm wide, leaf undivided part 50–90 µm (3–6 cells) high, cells in the lobe base 10–20 × 10–12 µm, cuticle distinctly papillose-verrucose, sometimes scarcely so (Korean-Japanese taxon) … T. pusilla
4. Taxonomic Treatment
Tetralophozia sibirica Vilnet et Bakalin sp. nov.
5. Materials and Methods
5.1. The Tetralophozia Overview
5.2. Specimens
5.3. Molecular-Genetic Study
5.4. Climate Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stephani, F. Species Hepaticarum; Georg: Geneva, Switzerland, 1909; Volume 3. [Google Scholar]
- Urmi, E. Tetralophozia Filiformis (Steph.) Comb. Nov. in Europa. J. Bryol. 1983, 12, 393–401. [Google Scholar] [CrossRef]
- Konstantinova, N.A. The Genus Tetralophozia (RM Schust.) Schljakov (Lophoziaceae, Hepaticae) in Russia. Arctoa 2002, 11, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Konstantinova, N.A.; Bakalin, V.A.; Potemkin, A.D.; Ignatov, M.S. Hepatic Flora of the upper Bureya River (Russian Far East). Arctoa 2002, 11, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Piippo, S. Annotated Catalogue of Chinese Hepaticae and Anthocerotae. J. Hattori Bot. Lab. 1990, 68, 1–192. [Google Scholar]
- Yamada, K.; Iwatsuki, Z. Catalog of the Hepatics of Japan. J. Hattori. Bot. Lab. 2006, 99, 1–106. [Google Scholar]
- Choi, S.-S.; Bakalin, V.A.; Sun, B.-Y. Unrecorded Liverwort Species from Mt. Deogyu, Korea. Korean J. Plant Taxon. 2010, 40, 218–225. [Google Scholar] [CrossRef]
- Vilnet, A.A.; Konstantinova, N.A.; Troitsky, A.V. Molecular Insight on Phylogeny and Systematics of the Lophoziaceae, Scapaniaceae, Gymnomitriaceae and Jungermanniaceae. Arctoa 2010, 19, 31–50. [Google Scholar] [CrossRef] [Green Version]
- Bakalin, V.A.; Vilnet, A.A.; Nguyen, V.S. Vietnamiella Epiphytica—A New Genus and Species of Anastrophyllaceae (Hepaticae). Bryologist 2020, 123, 48–63. [Google Scholar] [CrossRef]
- Bakalin, V.A. Liverworts of Stanovoye Nagorye Upland. Arctoa 2005, 13, 73–83. [Google Scholar]
- Damsholt, K.; Pugh, A. Illustrated Flora of Nordic Liverworts and Hornworts; Nordic Bryological Society Lund, 2002; ISBN 8798678124. [Google Scholar]
- Paton, J.A. The Liverwort Flora of the British Isles; Harley Books: Colchester, UK, 1999; ISBN 0-946589-60-7. [Google Scholar]
- Engel, J.J.; Merrill, G.L.S. Bonner’s “Types” Are Not Lectotypes. Nova Hedwig. 2019, 109, 63–64. [Google Scholar] [CrossRef]
- Schljakov, R.N. Novye Kombinacii i Novye Taksony Pechenochnyh Mkhov (Antocerotae i Hepaticae) [Combinationes et Taxa Anthocerotarum et Hepaticarum Nova]. Nov. Sist. Nizshih Rastenij 1976, 13, 225–229. [Google Scholar]
- Schuster, R.M. Notes on Nearctic Hepaticae. XIX. The Relationships of Blepharostoma, Temnoma and Lepicolea, with Description of Lophochaete and Chandonanthus Subg. Tetralophozia, Subg. n. J. Hattori Bot. Lab. 1960, 23, 192–210. [Google Scholar]
- Vána, J. Taxonomic Results of the BRYOTROP Expedition to Zaire and Rwanda 11. Cephaloziaceae, Cephaloziellaceae, Gymnomitriaceae, Jungermanniaceae, Lophoziaceae. Trop Bryol 1993, 8, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Schuster, R.M. Revisionary Studies of the Chandonanthoideae (Jungermanniales, Jungermanniaceae). Nova Hedwig. 2002, 74, 465–496. [Google Scholar] [CrossRef]
- Söderström, L.; Hagborg, A.; von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; Costa, D.P.; Crandall-Stotler, B.J.; et al. World Checklist of Hornworts and Liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef]
- Váňa, J.; Piippo, S. Bryophyte Flora of the Huon Peninsula, Papua New Guinea. XXXI. Cephaloziaceae Subfam. Alobielloideae, Cephaloziellaceae, Antheliaceae and Lophoziaceae (Hepaticae). Finn. Bot. Publ. Board 1989, 26, 263–290. [Google Scholar]
- Schuster, R.M. Austral Hepaticae. Part II. Nova Hedwig. 2002, 119, 1–606. [Google Scholar]
- Liu, Z.-W.; Peng, H. Notes on the Key Role of Stenochoric Endemic Plants in the Floristic Regionalization of Yunnan. Plant Divers 2016, 38, 289–294. [Google Scholar] [CrossRef]
- Inoue, H. Hepaticae and Anthocerotae of the Chichibu-Okutama Mountains, Central Japan. J. Hattori Bot. Lab. 1962, 25, 186–216. [Google Scholar]
- Takaki, N.; Amakawa, T.; Osada, T.; Sakuma, E. Bryophyte Flora of Mt. Kaikoma, Mt. Senjo and Mt. Kitadake (Southern Japan Alps). J. Hattori Bot. Lab. 1970, 33, 171–202. [Google Scholar]
- Feldberg, K.; Heinrichs, J.; Schmidt, A.R.; Váňa, J.; Schneider, H. Exploring the Impact of Fossil Constraints on the Divergence Time Estimates of Derived Liverworts. Plant Syst. Evol. 2013, 299, 585–601. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal Primers for Amplification of Three Non-Coding Regions of Chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.; Lickey, E.B.; Beck, J.T.; Farmer, S.B.; Liu, W.; Miller, J.; Siripun, K.C.; Winder, C.T.; Schilling, E.E.; Small, R.L. The Tortoise and the Hare II: Relative Utility of 21 Noncoding Chloroplast DNA Sequences for Phylogenetic Analysis. Am. J. Bot. 2005, 92, 142–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd., Oxford University Press: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Goloboff, P.A.; Catalano, S.A. TNT Version 1.5, including a Full Implementation of Phylogenetic Morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Keane, T.M.; Creevey, C.J.; Pentony, M.M.; Naughton, T.J.; Mclnerney, J.O. Assessment of Methods for Amino Acid Matrix Selection and Their Use on Empirical Data Shows That Ad Hoc Assumptions for Choice of Matrix Are Not Justified. BMC Evol. Biol. 2006, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum Likelihood-Based Phylogenetic Analyses with Thousands of Taxa and Mixed Models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Vilnet, A.A.; Klimova, K.G.; Ma, W.Z.; Choi, S.S.; Hentschel, J. Hidden in Plain View: An Example from Ptilidium (Ptilidiaceae, Marchantiophyta). Phytotaxa 2021, 510, 29–42. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Species | Infraspecific p-Distances, ITS1–2/trnL–F,% | Infrageneric p-Distances, ITS1–2/trnL–F,% | ||
|---|---|---|---|---|---|
| 1 | 2 | 3 | |||
| 1 | T. sibirica | 0.3/0.7 | |||
| 2 | T. filiformis | 1.1/0.0 | 2.5/1.2 | ||
| 3 | T. pusilla | 0.3/0.2 | 2.7/1.0 | 1.3/0.2 | |
| 4 | T. setiformis | 0.5/0.1 | 3.3/2.7 | 2.9/1.3 | 3.2/1.6 |
| № | Mark in Figure 2 | Accepted Name | Field no. | Latitude | Longitude | BIO01 | BIO02 | BIO03 | BIO04 | BIO05 | BIO06 | BIO07 | BIO08 | BIO09 | |
| 1 | square | T. filiformis | C-39-1-17 | 29.977055 | 101.88477 | 4.071 | 11.008 | 37.065 | 695.248 | 19.500 | −10.200 | 29.700 | 12.567 | −4.417 | |
| 2 | square | T. filiformis | C-40-16-17 | 29.976361 | 101.885194 | 4.071 | 11.008 | 37.065 | 695.248 | 19.500 | −10.200 | 29.700 | 12.567 | −4.417 | |
| 3 | square | T. filiformis | C-73-44-18 | 26.59494 | 99.764333 | 7.371 | 9.108 | 36.727 | 487.924 | 19.900 | −4.900 | 24.800 | 13.117 | 1.917 | |
| 4 | square | T. filiformis | V-10-6-19 | 22.505861 | 103.587805 | 9.813 | 7.758 | 39.382 | 420.606 | 19.600 | −0.100 | 19.700 | 14.717 | 5.017 | |
| 5 | triangle | T. pusila | 3735 | 35.664083 | 127.735583 | 9.108 | 10.767 | 29.660 | 923.811 | 26.700 | −9.600 | 36.300 | 20.017 | −2.733 | |
| 6 | triangle | T. pusila | 111058 | 35.333805 | 127.731972 | 5.796 | 8.608 | 26.487 | 867.395 | 21.900 | −10.600 | 32.500 | 15.900 | −2.683 | |
| 7 | triangle | T. pusilla | Kor-23-27-15 | 35.33166 | 127.73416 | 7.400 | 9.183 | 27.744 | 869.190 | 23.800 | −9.300 | 33.100 | 17.533 | −1.200 | |
| 8 | triangle | T. pusilla | Kor-25-1-15 | 35.43472 | 127.73083 | 11.188 | 10.958 | 29.941 | 929.507 | 29.400 | −7.200 | 36.600 | 22.183 | −0.667 | |
| 9 | triangle | T. pusilla | Kor-27-20-15 | 35.325 | 127.70694 | 6.475 | 8.900 | 27.052 | 868.830 | 22.800 | −10.100 | 32.900 | 16.583 | −2.083 | |
| 10 | triangle | T. pusilla | Kor-7-13-11 | 38.12777 | 128.44861 | 6.229 | 8.525 | 24.568 | 943.496 | 23.000 | −11.700 | 34.700 | 17.000 | −5.933 | |
| 11 | triangle | T. pusilla | J-88-40-15 | 35.74556 | 138.23389 | 2.033 | 8.900 | 27.900 | 837.942 | 18.400 | −13.500 | 31.900 | 12.300 | −7.983 | |
| 12 | circle | T. sibirica | MI-1077-97 | 52.06666 | 134.86666 | −8.296 | 15.075 | 27.409 | 1491.022 | 18.300 | −36.700 | 55.000 | 9.833 | −26.683 | |
| 13 | circle | T. sibirica | 37-11-00 | 56.90694 | 120.052305 | −7.938 | 12.008 | 22.279 | 1576.736 | 19.100 | −34.800 | 53.900 | 11.467 | −23.967 | |
| 14 | circle | T. sibirica | 13-24-01 | 51.42925 | 105.040583 | −0.975 | 9.800 | 23.113 | 1210.906 | 19.500 | −22.900 | 42.400 | 14.200 | −14.100 | |
| 15 | circle | T. sibirica | 411 | 56.911388 | 117.80944 | −9.108 | 12.483 | 24.193 | 1507.015 | 17.000 | −34.600 | 51.600 | 10.050 | −26.933 | |
| 16 | circle | T. sibirica | C. C. Exsiccata. 411 | 56.9115 | 117.8095 | −9.108 | 12.483 | 24.193 | 1507.015 | 17.000 | −34.600 | 51.600 | 10.050 | −26.933 | |
| 17 | snowflake | T. filiformis s.l. (Spain) | (Urmi. 1983) Urmi2028 (not seen) | 43.23194 | −1.52222 | 12.017 | 8.183 | 38.601 | 467.412 | 22.300 | 1.100 | 21.200 | 7.667 | 17.667 | |
| 18 | snowflake | T. filiformis s.l. (Spain) | (Urmi. 1983) Urmi 2220 (not seen) | 43.23333 | −1.53611 | 12.813 | 8.425 | 39.929 | 455.043 | 23.000 | 1.900 | 21.100 | 8.517 | 18.233 | |
| 19 | snowflake | T. filiformis s.l. | GBIF ALA B43214 (Alaska) | 56.160077 | −131.9698 | 6.183 | 7.117 | 32.057 | 547.309 | 16.400 | −5.800 | 22.200 | 3.083 | 11.350 | |
| 20 | snowflake | T. filiformis s.l. | GBIF ALA B43238 (Alaska) | 56.373755 | −132.100001 | 6.013 | 6.825 | 30.199 | 562.980 | 16.400 | −6.200 | 22.600 | 2.633 | 11.383 | |
| 21 | snowflake | T. filiformis s.l. | GBIF UBC B228679 (Canada) | 49.67 | −123.16 | 9.100 | 7.183 | 32.652 | 546.884 | 19.500 | −2.500 | 22.000 | 3.033 | 15.683 | |
| 22 | snowflake | T. filiformis s.l. | GBIF E BGBASE: 686287 (Bhutan) | 27.544167 | 90.722778 | 9.754 | 10.508 | 42.202 | 506.442 | 22.100 | −2.800 | 24.900 | 15.467 | 4.117 | |
| № | Mark in Figure 2 | Accepted Name | Field no. | Latitude | Longitude | BIO10 | BIO11 | BIO12 | BIO13 | BIO14 | BIO15 | BIO16 | BIO17 | BIO18 | BIO19 |
| 1 | square | T. filiformis | C-39-1-17 | 29.977055 | 101.88477 | 12.567 | −4.417 | 755.000 | 159.000 | 3.000 | 94.283 | 417.000 | 11.000 | 417.000 | 11.000 |
| 2 | square | T. filiformis | C-40-16-17 | 29.976361 | 101.885194 | 12.567 | −4.417 | 755.000 | 159.000 | 3.000 | 94.283 | 417.000 | 11.000 | 417.000 | 11.000 |
| 3 | square | T. filiformis | C-73-44-18 | 26.59494 | 99.764333 | 13.117 | 1.150 | 868.000 | 147.000 | 13.000 | 62.081 | 394.000 | 57.000 | 394.000 | 64.000 |
| 4 | square | T. filiformis | V-10-6-19 | 22.505861 | 103.587805 | 14.717 | 4.333 | 1756.000 | 367.000 | 5.000 | 87.810 | 967.000 | 40.000 | 967.000 | 71.000 |
| 5 | triangle | T. pusila | 3735 | 35.664083 | 127.735583 | 20.017 | −2.733 | 1510.000 | 345.000 | 30.000 | 81.675 | 833.000 | 110.000 | 833.000 | 110.000 |
| 6 | triangle | T. pusila | 111058 | 35.333805 | 127.731972 | 15.900 | −5.200 | 1833.000 | 402.000 | 36.000 | 77.820 | 987.000 | 135.000 | 987.000 | 139.000 |
| 7 | triangle | T. pusilla | Kor-23-27-15 | 35.33166 | 127.73416 | 17.533 | −3.650 | 1741.000 | 379.000 | 33.000 | 78.298 | 943.000 | 127.000 | 943.000 | 130.000 |
| 8 | triangle | T. pusilla | Kor-25-1-15 | 35.43472 | 127.73083 | 22.183 | −0.667 | 1401.000 | 311.000 | 24.000 | 82.863 | 779.000 | 94.000 | 779.000 | 94.000 |
| 9 | triangle | T. pusilla | Kor-27-20-15 | 35.325 | 127.70694 | 16.583 | −4.567 | 1784.000 | 392.000 | 35.000 | 78.138 | 963.000 | 132.000 | 963.000 | 134.000 |
| 10 | triangle | T. pusilla | Kor-7-13-11 | 38.12777 | 128.44861 | 17.133 | −5.933 | 1334.000 | 304.000 | 27.000 | 84.344 | 747.000 | 96.000 | 727.000 | 96.000 |
| 11 | triangle | T. pusilla | J-88-40-15 | 35.74556 | 138.23389 | 12.300 | −8.317 | 2066.000 | 325.000 | 46.000 | 58.026 | 900.000 | 167.000 | 900.000 | 239.000 |
| 12 | circle | T. sibirica | MI-1077-97 | 52.06666 | 134.86666 | 9.833 | −26.683 | 820.000 | 171.000 | 9.000 | 84.076 | 446.000 | 34.000 | 446.000 | 34.000 |
| 13 | circle | T. sibirica | 37-11-00 | 56.90694 | 120.052305 | 11.467 | −27.150 | 478.000 | 106.000 | 5.000 | 93.877 | 288.000 | 19.000 | 288.000 | 20.000 |
| 14 | circle | T. sibirica | 13-24-01 | 51.42925 | 105.040583 | 14.200 | −15.733 | 507.000 | 120.000 | 7.000 | 89.071 | 294.000 | 26.000 | 294.000 | 28.000 |
| 15 | circle | T. sibirica | 411 | 56.911388 | 117.80944 | 10.050 | −26.933 | 442.000 | 98.000 | 4.000 | 95.500 | 270.000 | 14.000 | 270.000 | 14.000 |
| 16 | circle | T. sibirica | C. C. Exsiccata. 411 | 56.9115 | 117.8095 | 10.050 | −26.933 | 442.000 | 98.000 | 4.000 | 95.500 | 270.000 | 14.000 | 270.000 | 14.000 |
| 17 | snowflake | T. filiformis s.l. (Spain) | (Urmi. 1983) Urmi 2028 (not seen) | 43.23194 | −1.52222 | 18.067 | 6.733 | 1241.000 | 134.000 | 66.000 | 21.016 | 377.000 | 226.000 | 233.000 | 348.000 |
| 18 | snowflake | T. filiformis s.l. (Spain) | (Urmi. 1983) Urmi 2220 (not seen) | 43.23333 | −1.53611 | 18.667 | 7.600 | 1232.000 | 136.000 | 65.000 | 21.606 | 377.000 | 223.000 | 235.000 | 344.000 |
| 19 | snowflake | T. filiformis s.l. | GBIF ALA B43214 (Alaska) | 56.160077 | −131.9698 | 12.967 | −0.283 | 2914.000 | 440.000 | 137.000 | 36.545 | 1071.000 | 444.000 | 470.000 | 832.000 |
| 20 | snowflake | T. filiformis s.l. | GBIF ALA B43238 (Alaska) | 56.373755 | −132.100001 | 12.967 | −0.717 | 2749.000 | 424.000 | 130.000 | 37.476 | 1016.000 | 421.000 | 447.000 | 780.000 |
| 21 | snowflake | T. filiformis s.l. | GBIF UBC B228679 (Canada) | 49.67 | −123.16 | 15.700 | 2.383 | 2692.000 | 470.000 | 68.000 | 60.630 | 1223.000 | 234.000 | 239.000 | 996.000 |
| 22 | snowflake | T. filiformis s.l. | GBIF E BGBASE: 686287 (Bhutan) | 27.544167 | 90.722778 | 15.467 | 3.150 | 974.000 | 211.000 | 5.000 | 87.628 | 537.000 | 21.000 | 537.000 | 24.000 |
| No | DCA | ||
|---|---|---|---|
| Axis 1 (X) | Axis 2 (Y) | Axis 3 (Z) | |
| 1 | 46.00 | 24.00 | 13.00 |
| 2 | 46.00 | 24.00 | 13.00 |
| 3 | 68.00 | 21.00 | 6.00 |
| 4 | 82.00 | 34.00 | 6.00 |
| 5 | 68.00 | 18.00 | 3.00 |
| 6 | 76.00 | 18.00 | 1.00 |
| 7 | 74.00 | 18.00 | 1.00 |
| 8 | 65.00 | 18.00 | 4.00 |
| 9 | 75.00 | 18.00 | 1.00 |
| 10 | 64.00 | 16.00 | 4.00 |
| 11 | 84.00 | 13.00 | 0.00 |
| 12 | 25.00 | 5.00 | 14.00 |
| 13 | 1.00 | 0.00 | 19.00 |
| 14 | 15.00 | 5.00 | 17.00 |
| 15 | 0.00 | 0.00 | 20.00 |
| 16 | 0.00 | 0.00 | 20.00 |
| 17 | 109.00 | 5.00 | 0.00 |
| 18 | 109.00 | 7.00 | 0.00 |
| 19 | 124.00 | 7.00 | 3.00 |
| 20 | 123.00 | 6.00 | 3.00 |
| 21 | 126.00 | 25.00 | 30.00 |
| 22 | 64.00 | 32.00 | 10.00 |
| Bioclimate Indices | Axis 1 (X) | Axis 2 (Y) | Axis 3 (Z) |
|---|---|---|---|
| BIO01 | 0.803697855 | 0.610695077 | −0.577801558 |
| BIO02 | −0.843076109 | −0.290005672 | 0.423780285 |
| BIO03 | 0.505493332 | 0.586507785 | −0.228884513 |
| BIO04 | −0.843868158 | −0.616066578 | 0.463700903 |
| BIO05 | 0.14350037 | 0.347252607 | −0.4291601 |
| BIO06 | 0.849729356 | 0.6223567 | −0.536279158 |
| BIO07 | −0.873734551 | −0.569422916 | 0.452828573 |
| BIO08 | −0.348310994 | 0.390696387 | −0.310065311 |
| BIO09 | 0.92838766 | 0.418628246 | −0.450694567 |
| BIO10 | 0.475267305 | 0.344690362 | −0.51527616 |
| BIO11 | 0.846435514 | 0.614736776 | −0.540815429 |
| BIO12 | 0.858486569 | 0.221188912 | −0.359161055 |
| BIO13 | 0.689773053 | 0.400419913 | −0.330532825 |
| BIO14 | 0.795660002 | −0.224500064 | −0.359993131 |
| BIO15 | −0.79457789 | 0.224928435 | 0.473925358 |
| BIO16 | 0.703195812 | 0.421098664 | −0.315688842 |
| BIO17 | 0.821092389 | −0.195327474 | −0.390334625 |
| BIO18 | 0.189870135 | 0.48707667 | −0.596398852 |
| BIO19 | 0.778554616 | −0.068321891 | 0.013610967 |
| No | Name | Label Data | Latitude. N | Longitude. E | Field Number Plus Barcode and Herbarium Acronym (in Brackets) | GenBank Accession Number | |
|---|---|---|---|---|---|---|---|
| ITS1–2 nrDNA | trnL–F/trnG–intron cpDNA | ||||||
| 1 | Tetralophozia filiformis | China. Sichuan Province. Bakalin & Klimova. 13 October 2017 | 29.977055 | 101.88477 | China-39-1-17 (VBGI- 37281. KPABG-122599 duplicate) | MZ231275 | MZ229433/- |
| 2 | T. filiformis | China. Sichuan Province. Bakalin & Klimova. 13 October 2017 | 29.976361 | 101.885194 | China-40-16-17 (VBGI-37325) | no data | no data |
| 3 | T. filiformis | China. Yunnan Province. Bakalin & Ma. 11 October 2018 near locus classicus of the species | 26.59494 | 99.764333 | C-73-44-18 (VBGI) | MZ231276 | MZ229434/- |
| 4 | T. filiformis | Vietnam. Lai Châu Province. Bakalin & Klimova | 22.505861 | 103.587805 | V-10-6-19 (VBGI-65792) | no data | no data |
| 5 | T. pusilla | Republic of Korea Gyeongsang-do. Choi. 14 June 2009 | 35.664083 | 127.735583 | Choi-3735 (JNU. duplicate VBGI) | no data | no data |
| 6 | T. pusilla | Republic of Korea Gyeongsang-do. Choi. 1 October 2011 | 35.333805 | 127.731972 | Choi-111058 (JNU. duplicate VBGI) | no data | no data |
| 7 | T. pusilla | Republic of Korea. Gyeongsangnam-do. Bakalin. 5 May 2015 | 35.33166 | 127.73416 | Kor-23-27-15 (VBGI) | no data | no data |
| 8 | T. pusilla | Republic of Korea. Gyeongsangnam-do. Bakalin. 6 May 2015 | 35.43472 | 127.73083 | Kor-25-1-15 (VBGI) | no data | no data |
| 9 | T. pusilla | Republic of Korea. Gyeongsangnam-do. Bakalin 7 May 2015 | 35.32500 | 127.70694 | Kor-27-20-15 (VBGI). (KPABG-120508. duplicate) | MZ231277 | MZ229435/- |
| 10 | T. pusilla | Republic of Korea. Gangwon-do. Bakalin. 11 May 2015 | 38.12777 | 128.44861 | Kor-7-13-11 (VBGI) | no data | no data |
| 11 | T. pusilla | Japan. Yamanashi Prefecture. Bakalin. 1 October 2015 near locus classicus of the species | 35.74556 | 138.23389 | J-88-40-15 (VBGI-5796. KPABG-123441 duplicate) | MZ231278 | MZ229436/MZ229442 |
| 12 | T. sibirica | Russia. Khabarovsk Territory. Ignatov. 15 August 1997 | 52.06666 | 134.86666 | MI-1077-97 (KPABG-116740) | no data | MZ229438/- |
| 13 | T. sibirica | Russia. Amurskaya Province. Bakalin. 17 August 2000 | 56.90694 | 120.052305 | 37-11-00 (KPABG-101730) | no data | MZ229439/- |
| 14 | T. sibirica | Russia. Buryatia Republic. Konstantinova. 4 August 2001 | 51.42925 | 105.040583 | 13-24-01 (KPABG-102424) | EU791792 | EU791669/- |
| 15 | T. sibirica | Russia. Zabaikalsky Territory. Mamontov. 7 July 2013 | 56.911388 | 117.80944 | 411 (KPABG-121349) | MZ231279 | MZ229437/- |
| 16 | T. setiformis | Russia: Buryatia Republic. Konstantinova & Savchenko. 8 August 2002 | 51.185591 | 105.181264 | 123-2-02 (KPABG-121659) | EU791793 | EU791670/- |
| 17 | T. setiformis | Russia: Kamchatka Territory. Bakalin. 13 July 2006 | 55.901388 | 158.782777 | 99-06 (KPABG-112052) | MZ231281 | MZ229441/- |
| 18 | T. setiformis | Russia: Murmansk Province. Konstantinova. 7 July 2007 | 67.3229 | 35.1623 | K201-1-07 (KPABG-18022) | MZ231280 | MZ229440/- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalin, V.A.; Vilnet, A.A.; Maltseva, Y.D.; Klimova, K.G.; Bakalin, D.A.; Choi, S.S. Hidden Diversity within Tetralophozia filiformis (Marchantiophyta, Anastrophyllaceae) in East Asia. Plants 2022, 11, 3121. https://doi.org/10.3390/plants11223121
Bakalin VA, Vilnet AA, Maltseva YD, Klimova KG, Bakalin DA, Choi SS. Hidden Diversity within Tetralophozia filiformis (Marchantiophyta, Anastrophyllaceae) in East Asia. Plants. 2022; 11(22):3121. https://doi.org/10.3390/plants11223121
Chicago/Turabian StyleBakalin, Vadim A., Anna A. Vilnet, Yulia D. Maltseva, Ksenia G. Klimova, Daniil A. Bakalin, and Seung Se Choi. 2022. "Hidden Diversity within Tetralophozia filiformis (Marchantiophyta, Anastrophyllaceae) in East Asia" Plants 11, no. 22: 3121. https://doi.org/10.3390/plants11223121