Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue


1
Department of Botany and Zoology, Biosciences Center, Federal University of Rio Grande do Norte (UFRN), Highway BR-101, Lagoa Nova, Campus Universitario, Natal 59072-970, RN, Brazil
2
Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, Pl-31-016 Kraków, Poland
3
Institut National De La Recherche Agronomique Clermont-Auvergne-Rhônes-Alpe, INRAE, 63000 Clermont-Ferrand, France
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(2), 221; https://doi.org/10.3390/d15020221
Submission received: 28 January 2023
/
Accepted: 1 February 2023
/
Published: 3 February 2023
(This article belongs to the Special Issue Systematics, Ecology and Taxonomy of Collembola)
{kind=link}
{kind=link}
{kind=link}
Springtails (Collembola) are very small terrestrial arthropods commonly found to be associated with edaphic environments. Since most species range from 1 to 5 mm and live hidden under rocks and plant debris, or even inside the soil [1], they are unknown to most people, including some agricultural workers. The tiniest recorded springtail, Sphaeridia pilleata Bretfeld and Gauer, 1994 from Brazil, is arguably the smallest known adult hexapod, with its smaller males measuring about 0.12 mm [2,3].
Collembola is placed within Hexapoda, among its basal lineages. In the past, Hexapoda and Insecta were considered synonyms by different authors, but the current systematics of Arthropoda supports Insecta as an independent class of Hexapoda subphylum, with Collembola, Protura and Diplura as the other non-insect classes [4,5]. The main feature used to separate the latter three classes from Insecta is the entognathy, a condition in which the mouthparts are hidden inside the head capsule. The insects, on the other hand, are ectognathous, and their mandibles and/or maxillae are often very well developed and exposed [6].
Springtails tagmosis is remarkably similar to the one seen in Insecta. Their body is divided into a head, thorax and abdomen. The head always bears a pair of anterior four-segmented antennae, but in some taxa there can be subsegmentation or partial fusions of the antennal segments. The thorax has three segments similarly to other hexapods, pro, meso and metathorax, and each one has a pair of legs. The number of abdominal segments is fixed in Collembola, and it is always six, although in different lineages the fusion of segments may obscure this observation (Figure 1A) [1]. Springtails morphologically diverge from the other hexapod classes especially by their exclusive abdominal appendages [4] (Figure 1A). The most evident is the jumping organ, the furca (or furcula), which gave them their common name. This appendage is located in the ventral face of the fourth abdominal segment and it is primarily used to escape from predators. The furca works in synergy with another abdominal appendage, the tenaculum (or retinaculum). This small structure is found in the ventral side of the third abdominal segment and has some apical teeth which hold the furca rested near the body. Whenever in danger, these appendages are detached from each other, the furca is quickly pressed against the ground and the springtail jumps (Figure 1B,C). However, in many euedaphic taxa, both furca and tenaculum may be strongly reduced or entirely missing [7]. So, a better synapomorphy of springtails to circumscribe the group is the ventral tube (or collophore), which is also located in the ventral face of the abdomen (in the first segment). This very peculiar appendage holds two apical vesicles which are primarily used in the fluid balance but can also help in the oxygen absorption, grooming, jumping and attaching the specimens to smooth surfaces [1,7]. The ventral tube is present in all sprigtails and was used by Sir John Lubbock in 1870 to name the class as “Collembola” (from Latin “colla” = glue; from Greek “embolon” = a plug) [1].
Similar to the Insecta, the largest internal lineages of Collembola are considered as orders, and currently there are four of them: Entomobryomorpha, Poduromorpha, Symphypleona and Neelipleona [1]. The Entomobryomorpha and Poduromorpha hold the taxa with clearly segmented trunks, and the main difference between both is the presence of the pronotum (the dorsal plate of the prothorax) in the latter, which is missing in the Entomobryomorpha (Figure 1A and Figure 2A–D). These two orders may also be separated in most of the cases by other morphological traits: adult Entomobryomorpha usually have more abundant and/or heterogeneous body chaetae, especially on the dorsal trunk and legs, and longer appendages (antennae, legs, ventral tube and furca), but exceptions are seen among species which live inside the soil [1]. The Symphypleona and Neelipleona (Figure 2E–H) are represented by species with most trunk segments fused to each other, resulting in a globular body. The Neelipleona are remarkably different from the Symphypleona by their reduced antennae, shorter than the head, the largest portion of their trunk formed by enlarged thoracic segments, the presence of long coxae and subsegmented dens. Contrastingly, the Symphypleona have antennae which are at least longer than the head, sometimes even longer than the body, a trunk mostly constituted by abdominal segments, short coxae and unsegmented dens [3] (Figure 2E–H).
The oldest fossil of Collembola is Rhyniella praecursor Hirst and Maulik, 1926 from the Early Devonian (ca 400 Ma), which is also the earliest true hexapod recorded so far, and one of the oldest known evidence of a terrestrial metazoan [1,10]. Even so, the species matches the morphology of some contemporary taxa, being placed in the extant family Isotomidae [11]. Actually, the remains from the Rhynie chert may actually include hypogastrurid and neanurid taxa as well, probably representing five species (personal communication). These data strongly support that the Collembola origin is quite older, and at least some of its main lineages, such as extant orders, superfamilies and families, were already established during the Early Devonian.
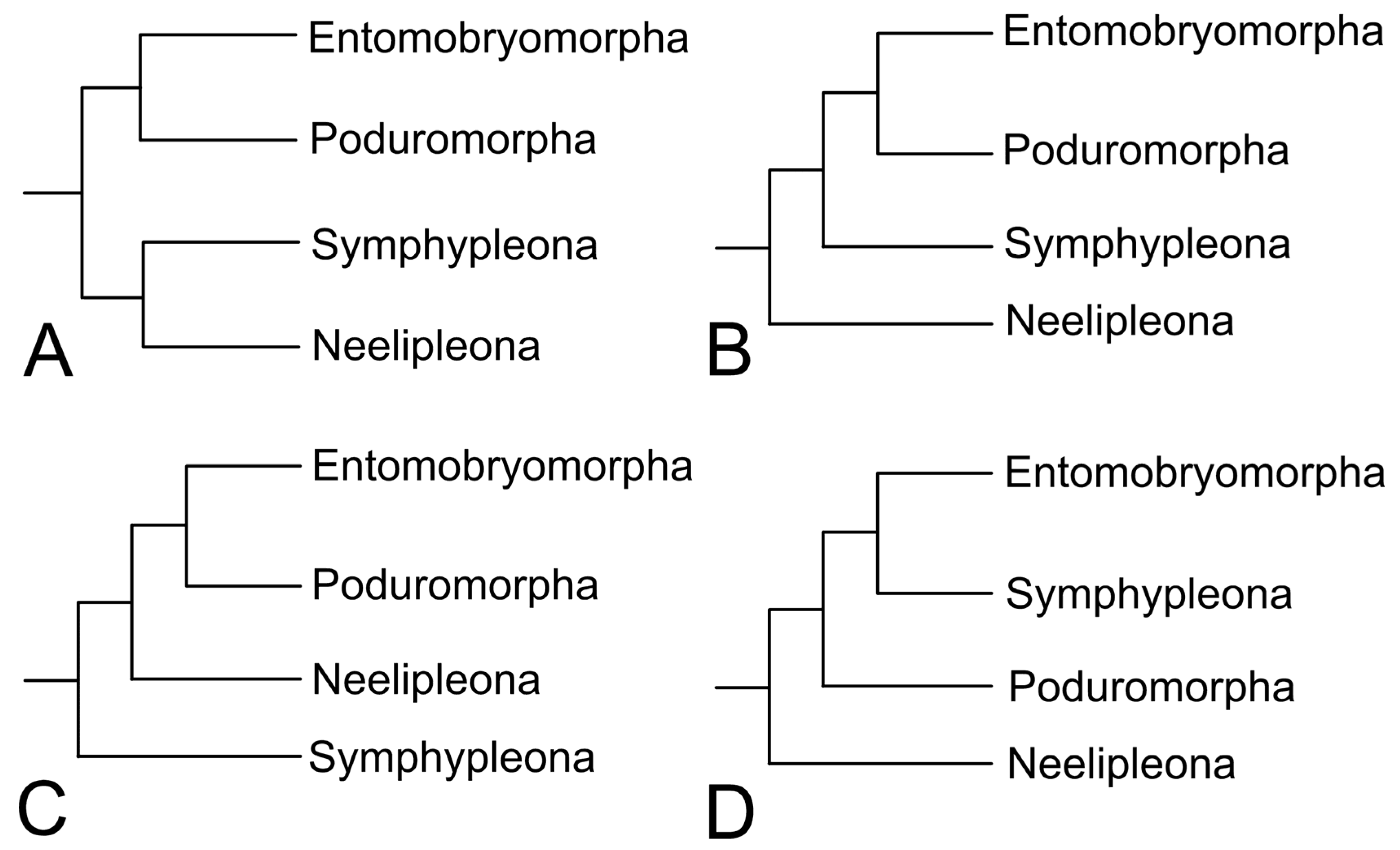
Considering the internal evolution of springtails and its systematics, many phylogenetic studies and hypotheses were published during the last century, based on internal and external morphology and molecular markers [9]. The systematics of the familiar and generic levels of springtails has greatly advanced in recent years, especially of the Entomobryomorpha, and changes have been constantly published, based mostly on phylogenetic grounds [12,13,14,15,16,17,18,19]. Some of these studies are listed and discussed in one paper of this Special Issue [9]. Possibly the most puzzling debate regarding this topic is the relationship of the current four orders and many topologies were proposed in order to relate them (Figure 3). Since these lineages are remarkably ancient and show many autapomorphies, very few morphological traits are considered reliable to investigate the orders relationships, if any. Likewise, the use of different molecular markers, datasets and types of analyses reached very dissonant results so far [9] (Figure 3). Perhaps, and unfortunately in an anticlimactic way, the only consensus on this issue is that further studies are in need of clearly unveiling the internal evolution of Collembola main lineages.
The first formal taxonomic description of a springtail species was possibly provided by De Geer in 1743, while the group was also the subject of Linnaeus studies, summarized in his influential work, the 10th edition of the “Systema Naturae”, as noted by Hopkin [7]. Now, there are about 9400 nominal extant species of springtails, with more than half of them described during the last 50 years. Most of these species were recorded and described from the Holarctic region, especially due to the efforts of many important European and North American researchers [1]. However, a large number of species have been described during the last decades from the Neotropical, Oriental and Australian regions. At the same time, the tropical ecosystems are likely to shelter the highest number of undescribed species. For instance, in the past two decades, the number of recorded species from Brazil more than doubled, and organizing part of this data was the subject of one study of our Special Issue [24]. As another example of the expansion of the knowledge on the Neotropical fauna, a new genus and species from Brazil are described in one more paper of this Collembola volume [25]. In this scenario, we are possibly witnessing a “latitudinal” inversion of the global taxonomic efforts to uncover and describe this fauna [26]. Different estimates suggest something between 50.000 and 500.000 living species of Collembola [7,27] and, in any case, we are very far from recognizing the majority of them. The higher numbers of such projections take into account the cryptic diversity, which were already recorded within many taxa of springtails, as in [27]. This subject is the main theme of another paper of our Special Issue [28].
Springtails are widely distributed in terrestrial environments, but they can also occur on the surface of standing puddles of fresh or salt water, or even on underground littoral waters [29]. Collembola are recorded from all continents, including the high Arctic and Antarctica, and can live in arid desert areas to snow fields and permanent ice [7,29]. Most species, however, live associate to the soil, and in this environment, they can be categorized as: epedaphic (or epigeic—species which inhabit the aboveground, including the litter layer), hemiedaphic (taxa which can live both within the soil and on its surface) and euedaphic (species which only live inside the soil). Some euedaphic species may penetrate the soil more than 1.5 m below the surface, although most of them live on shallower depths [7,30]. In the edaphic environments, some species may also occur within insect nests, such as those of ants (myrmecophily) and termites (termitophily) [1]. Taxa linked to the vegetation are called atmobiotic and can live associated with grasses and bushes to the forest canopy, being occasionally also found on the litter layer [31]. Many springtail species occur inside caves, with some of them strongly adapted to their dark zones. Such species may have a reduction in or entirely lack body pigments and eyes and may present enlarged appendages or other troglomorphic traits [32]. In some cases, such species share a long evolutionary history with the underground environments, to the point where their distribution is restricted to one or few connected cavities (troglobites).
The population sizes of springtails can greatly vary from one ecosystem to the other, but their densities in most terrestrial environments range from 104 to 105 individuals m−2. Even so, higher numbers were recorded from different countries and biomes [7]. In number of specimens, their densities may only be surpassed by the Acari, but in some habitats they can represent more than half of the total soil arthropods individuals belonging to the mesofauna [1,7].
Collembola density may strongly influence the soil functioning and ecosystem services delivery. The fecal pellets and urine of Collembola, mainly from hemi and euedaphic species, may represent a large part of the humus and base nutrient, increasing the soil fertility and favoring plant growth [33,34,35]. Collembola not only improve these soil and plant properties but they can also alter the nutrient distribution in the soil and, consequently, the functional characteristics of the plants related to the nutrient uptake, especially the root architecture. Some studies showed that when the abundance of euedaphic species is high in the rhizosphere, plants reduce root growth due to the high nutrient concentration close to them and may direct that extra energy to improve shoot traits such as the biomass, flowering phenology and seed production [36,37]. Such a pattern is mainly observed in the presence of high densities of euedaphic springtails, possibly due to their feeding activities and their fecal pellet and urine production being limited to the roots zone.
Although Collembola species support complex food chains involving small predators such as diplurans, staphylinid and carabid beetles, ants, centipedes, harvestmen, mites, spiders and pseudoscorpions [1,7,35], they are well known for regulating other communities by feeding on a wide range of microorganisms (fungi, bacteria and actinomycetes). In fact, the best studied mechanism in which Collembola affect soil functioning is through the regulation of the soil microbial community. By feeding mainly on fungi, especially senescent hyphae and spores, springtails control the size of the fungal population and stimulate their growth, changing fungi–plant interactions. For instance, there is evidence that Collembola may regulate the communities of pathogenic and symbiotic microorganisms (like mycorrhizae), which in turn can influence plant growth and primary productivity [7,30,34,35]. Collembola can also feed on a wide range of non-microbial sources, such as plant debris, algae, pollen, decaying animals and feces [7,34,38]. There are even some reports of phytophagous species which feed on live plant tissues, turning few taxa such as Sminthurus viridis (Linnæus, 1758) into occasional agricultural pests [7]. Rarely, springtails can be carnivorous or cannibalistic, and such feeding preferences may play an important role in extreme oligotrophic conditions, such as in cave ecosystems [29,39].
Collembola feeding activity usually contributes to 1–10% of soil respiration, but in particular ecosystems, such a contribution can raise until 37% [7,30,35]. This activity is an important driver of multiple ecosystem functions related to the soil biochemical processes, such as decomposition, carbon stock, nutrient cycling, and plant nutrition. Additionally, springtails may shift edaphic physical properties by contributing to soil aggregate formation, either by stimulating the growth of fungal hyphae or by producing secondary compounds which act in the entanglement of microaggregates and macroaggregate stabilization [40]. The main results of these processes are the increase in the soil porosity, which influences the gas and water exchanges efficacy between the soil and atmosphere, and edaphic stability against the erosion process [40]. The fragmentation of plant debris and the production of feces also change the soil’s physical structure [34,35]. Finally, the horizontal and vertical mobility of some species of Collembola, especially of hemi and epedaphic taxa, contributes to the dispersion of spores and other microbial cells [7]. Therefore, in addition to altering the composition of species in a given local niche, springtails can contribute significantly to the colonization of new environments, probably being important to the recovery of degraded environments.
Collembola diversity and distribution is largely affected by environmental variables. At a regional scale, climate plays an important role since it filters select species adapted to different conditions. Most Collembola depend on moist habitats under certain temperatures [35], but there are species adapted to drought, extreme cold, the total absence of light, among other limiting conditions. At a local scale, the soil physical–chemical properties, and vegetation structure and composition are major drivers of Collembola assembly, since they provide shelter, shading and food resources to most species [7,30,41]. Additionally, guilds of predators may regulate springtails communities, affecting the species distribution and population sizes [35]. Conceivably, environmental changes, especially those caused by anthropogenic actions, affect Collembola diversity and distribution. These changes may affect the structure and composition of springtails communities and influence all the related functions delivered by their activities. Among the main threats for soil fauna, including Collembola, we can cite the native habitats conversion into crops, artificial forestry or livestock farms; the urban and industrial expansion; soil and water pollution; and the introduction of invasive alien species [35]. One study published in our Special Issue exemplifies the negative effects of native grassland conversion into Eucalyptus plantation on the Collembola community in South Brazil [41], as explained in the next topic.
- Diversity’s Special Issue: Systematics, Ecology and Taxonomy of Collembola
In this Special Issue of Diversity, we invited authors to publish relevant studies on some of the main subjects regarding the Collembola knowledge: systematics, ecology and taxonomy. The five papers of this issue addressed, to some extent, all of these topics.
Regarding the taxonomy and species richness, three papers [24,25,28] provided important contributions to the understanding of taxa distribution, delimitation, morphology and cryptic species diversity in different aspects. In Lima et al. [24], a detailed checklist of Brazilian Entomobryomorpha is presented, including species distribution, new records to Brazilian oceanic islands and the detailed redescription of Seira musarum Ridley, 1890, which represents the first record and species description from Brazil. The survey and summarization of such data is rather important, especially due to the fast increase in the number of registered and described species of Collembola in the country. For instance, there was 199 species of springtails recorded from Brazil in 2003, with 91 of them belonging to Entomobryomorpha [42]. Now, only 20 years later, these numbers increased to 475 and 247, respectively [24,43]. The data on the endemic insular fauna provided by the authors are particularly relevant to the conservation, since some of these populations may have very limited distributions, are isolated and are thus more vulnerable to environmental changes.
In the second taxonomical study [25], a new genus and species of Sminthurididae are described from Brazil and compared to other taxa of the family. Sminthurididae is one the most fascinating Collembola families due to the species’ courtship behavior and males’ secondary sexual dimorphism. In this lineage, males are substantially smaller than the females and have highly modified antennal claspers used to hold on to the opposite sex before mating [3,44] (Figure 2F). The Sminthurididae are quite diverse in the Neotropical Region [1,25], but the specimens’ small size and morphological complexity may impose difficulties in the taxonomic study and description of this particular fauna. In Medeiros et al. [25], the authors described in detail a new genus, Parasminthurides Medeiros and Bellini, 2022, and a new species, P. spinosus Medeiros and Bellini, 2022, providing a detailed view of both sexes’ morphology. The authors also surveyed the most relevant diagnostic characters of all the genera in the family, presenting a comparative plate of male’s antennal claspers of all Sminthurididae genera and an identification key to them. In the end of the paper, there is also a discussion on the past and current systematics of Sminthurididae, indicating the family and the Symphypleona as a whole should be better investigated in the light of phylogenetics.
The third taxonomical paper of our Special Issue, Dukes et al. [28], investigated the occurrence and intraspecific diversity of Symphypleona and Neelipleona taxa from the southern high Appalachia (USA). The region shelters a rich and partially endemic biota, but it is still understudied concerning the springtails [28]. The authors used COI metabarcoding techniques to investigate the Collembola species and morphospecies of the above orders and found some very interesting results. While the morphology supported 25 to 30 sampled species, different automated taxa delimitation methods found a remarkable intraspecific diversity, supporting 43 to 90 putative species. These data strongly suggest some isolate populations may represent cryptic taxa. On the other hand, the use of DNA barcodes to identify Symphypleona and Neelipleona failed, pointing to a lack of representative and reliable molecular information available on public databases.
An ecological perspective of the impacts of the conversion of natural habitats into crops in the epigeic Collembola communities was the subject of Silva et al.’s [41] work. The study was conducted in the Brazilian Pampa, a species-rich Neotropical phytogeographic domain mainly covered by grasslands, found in the southern region of the country [41]. The authors investigated and compared two different types of land-use: native grasslands used to livestock activity and Eucalyptus plantation. The artificial afforestation significantly affected the native Collembola communities, strongly reducing their abundance, species richness and alpha diversity. Such results had as the main driving factors the decrease in plant richness and changes in the plant dominance in the cropping fields. Species turnover (replacement) better explained the differences in springtails species composition, supporting that the conversion of native grasslands into Eucalyptus caused local extinctions, selecting a few species better adapted to the artificial conditions. At the same time, the results also pointed out the Brazilian Pampa holds a diverse and still mostly unknown fauna of epigeic springtails, which is already highly endangered by different types of land-use [41,43].
The final paper of our volume investigated the evolution of Collembola higher taxa based on mitogenomes [9]. Such markers were already successfully used to investigate the evolution and systematics of many animal lineages, including Collembola. However, molecular phylogenies based on different markers provided very discrepant results compared to each other and to traditional hypotheses during the recent years [9]. In this paper, a large-scale phylogenetic investigation of the main clades of Collembola was performed based on amino acid sequences of mitogenomes of 124 species. Such effort resulted in one of the most representative phylogenies of springtails published so far, which endorsed many traditional and more recent hypotheses discussed in the paper. For instance, the main tree supported the validity of the four orders, the superorder Arthropleona (Entomobryomorpha + Poduromorpha), the sister-group relationship of the Entomobryoidea with the Isotomidae, the Neanuroidea, with Brachystomellidae as one of the internal branches of Neanuridae, and the suborders of Symphypleona, Appendiciphora and Sminthuridida. The results of this study also supported the grouping of Neelidae genera into two subfamilies: Neelinae and Neelidinae. As expected, the results of this analysis also dismissed some controversial groups such as the superorder Symphypleona s. lat. (Symphypleona + Neelipleona) and Paronellidae, supported Hypogastruridae as a paraphyletic taxon and the need of a revision of Isotomidae subfamilies [9].
- Knowing More about Springtails
In this brief paper, we listed some key facts of Collembola and the main findings of the papers published in Diversity’s Collembola special volume. If you are looking for more general data on springtails, we suggest starting with the “Checklist of the Collembola of the World” database [1], which is kept constantly updated by Dr. Frans Janssens. This website gathers a large amount of relevant data regarding springtails study history, morphology, overall biology, ecology, phylogeny, systematics, identification keys to many taxa in different taxonomical levels, plus the most comprehensive checklist of Collembola currently available. Similarly, Hopkin’s book “Biology of the Springtails (Insecta: Collembola)” [7] is one of the most important general data compilations concerning the group published so far. Further general data on the Collembola is also presented in Christiansen [29], Christiansen and Bellinger [45], Rusek [34] and Potapov et al. [35].
Books, monographs and dedicated papers regarding the morphology, systematics and taxonomy of Collembola, including genera and species identification, are also available, especially for the Palaearctic fauna. Concerning the Poduromorpha, we suggest Stach [46,47,48,49], Massoud [50], Dunger [51], Cassagnau [52], Weiner [53], Najt and Weiner [54], Jordana et al. [55], Pomorski [56], Fjellberg [57], Thibaud et al. [58], Weiner and Najt [59], Najt et al. [60] and Paśnik and Weiner [61]; for the Entomobryomorpha, Stach [62,63,64], Potapov [65], Fjellberg [66] and Jordana [67]; for the Symphypleona, Stach [68,69], Richards [70], Betsch [44], Bretfeld [3] and Fjellberg [66]; and for the Neelipleona, Bonet [71], Stach [69] and Schneider [72]. Data on the North American taxa of all four orders, including the Hawaiian fauna, are available in Christiansen and Bellinger [45,73].
Acknowledgments
We would like to thank Gleyce Medeiros for revising the first draft of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 24 January 2023).
- Bretfeld, G.; Gauer, U. Diagnostic description of the males of new Sphaeridia species (Insecta, Collembola) from South America. Andrias 1994, 13, 113–136. [Google Scholar]
- Bretfeld, G. Synopses on Palaeartic Collembola, Volume 2. Symphypleona. Abh. Ber. Naturkundemus. Gorlitz 1999, 71, 1–318. [Google Scholar]
- Grimaldi, D.; Engel, M.S. Evolution of the Insects, 1st ed.; Cambridge University Press: New York, NY, USA, 2005; pp. 1–755. [Google Scholar]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Beutel, R.G.; Yavorskaya, M.I.; Mashimo, Y.; Fukui, M.; Meusemann, K. The Phylogeny of Hexapoda (Arthropoda) and the Evolution of Megadiversity. Proc. Arthropod. Embryol. Soc. Jpn. 2017, 51, 1–15. [Google Scholar]
- Hopkin, S.P. Biology of the Springtails (Insecta: Collembola), 1st ed.; Oxford University Press: New York, NY, USA, 1997; pp. 1–330. [Google Scholar]
- Bellini, B.C. Colêmbolos: Uma riqueza microscópica no Semiárido. In Conhecendo os Artrópodes do Semiárido, 1st ed.; Bravo, F., Calor, A.R., Eds.; Métis Produção Editorial: São Paulo, Brazil, 2016; Volume 1, pp. 43–55. [Google Scholar]
- Bellini, B.C.; Zhang, F.; Souza, P.G.C.; Santos-Costa, R.C.; Medeiros, G.S.; Godeiro, N.N. The Evolution of Collembola Higher Taxa (Arthropoda, Hexapoda) Based on Mitogenome Data. Diversity 2023, 15, 7. [Google Scholar] [CrossRef]
- Davis, R.B.; Baldauf, S.L.; Mayhew, P.J. Many hexapod groups originated earlier and withstood extinction events better than previously realized: Inferences from supertrees. Proc. R. Soc. B Biol. Sci. 2010, 277, 1597–1606. [Google Scholar] [CrossRef]
- Greenslade, P.; Whalley, P.E.S. The systematic position of Rhyniella praecursor Hirst & Maulik (Collembola). The earliest known hexapod. In Proceedings of the Second International Seminar on Apterygota, Siena, Italy, 4–6 September 1986; Volume 1, pp. 319–323. [Google Scholar]
- Soto-Adames, F.N.; Barra, J.A.; Christiansen, K.; Jordana, R. Suprageneric classification of Collembola Entomobryomorpha. Ann. Entomol. Soc. Am. 2008, 101, 501–513. [Google Scholar] [CrossRef]
- Zhang, F.; Deharveng, L. Systematic revision of Entomobryidae (Collembola) by integrating molecular and new morphological evidence. Zool. Scr. 2015, 44, 298–311. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, D.; Yu, D.; Wang, B. Molecular phylogeny supports S-chaetae as a key character better than jumping organs and body scales in classification of Entomobryoidea (Collembola). Sci. Rep. 2015, 5, 12471. [Google Scholar] [CrossRef]
- Zhang, F.; Bellini, B.C.; Soto-Adames, F.N. New insights into the systematics of Entomobryoidea (Collembola: Entomobryomorpha): First instar chaetotaxy, homology and classification. Zool. Syst. 2019, 44, 249–278. [Google Scholar] [CrossRef]
- Zhang, F.; Cipola, N.G.; Pan, Z.-X.; Ding, Y. New insight into the systematics of Heteromurini (Collembola: Entomobryidae: Heteromurinae) with special reference to Alloscopus and Sinodicranocentrus gen. n. Arth. Syst. Phylogeny 2020, 78, 1–16. [Google Scholar] [CrossRef]
- Godeiro, N.N.; Pacheco, G.; Liu, S.; Cipola, N.G.; Berbel-Filho, W.M.; Zhang, F.; Gilbert, M.T.; Bellini, B.C. Phylogeny of Neotropical Seirinae (Collembola, Entomobryidae) based on mitochondrial genomes. Zool. Scr. 2020, 49, 329–339. [Google Scholar] [CrossRef]
- Godeiro, N.N.; Ding, Y.; Cipola, N.G.; Jantarit, S.; Bellini, B.C.; Zhang, F. Phylogenomics and systematics of Entomobryoidea (Collembola): Marker design, phylogeny and classification. Cladistics 2022, 1–15. [Google Scholar] [CrossRef]
- Yu, D.; Ding, Y.; Tihelka, E.; Cai, C.; Hu, F.; Liu, M.; Zhang, F. Phylogenomics of Elongate-Bodied Springtails Reveals Independent Transitions from Aboveground to Belowground Habitats in Deep Time. Syst. Biol. 2022, 71, 1023–1031. [Google Scholar] [CrossRef]
- Gao, Y.; Bu, Y.; Luan, Y.-X. Phylogenetic Relationships of Basal Hexapods Reconstructed from Nearly Complete 18S and 28S rRNA Gene Sequences. Zool. Sci. 2008, 25, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Cruaud, C.; D’Haese, C.A. Unexpected diversity in Neelipleona revealed by molecular phylogeny approach (Hexapoda, Collembola). Soil Org. 2011, 83, 383–398. [Google Scholar]
- Schneider, C.; D’Haese, C.A. Morphological and molecular insights on Megalothorax: The largest Neelipleona genus revisited (Collembola). Invertebr. Syst. 2013, 27, 317–364. [Google Scholar] [CrossRef]
- Leo, C.; Carapelli, A.; Cicconardi, F.; Frati, F.; Nardi, F. Mitochondrial genome diversity in Collembola: Phylogeny, dating and gene order. Diversity 2019, 11, 169. [Google Scholar] [CrossRef]
- Lima, E.C.A.; Lopes, B.C.H.; Oliveira-Neto, M.A.; Mendonça, M.C.; Zeppelini, D. Synthesis of Brazilian Entomobryomorpha (Collembola: Hexapoda) with Special Emphasis on the Equatorial Oceanic Islands and Redescription of the First Species of Collembola Recorded in Brazil. Diversity 2022, 14, 553. [Google Scholar] [CrossRef]
- Medeiros, G.S.; Nunes, R.C.; Zhang, F.; Godeiro, N.N.; Bellini, B.C. A New Genus of Sminthurididae (Collembola, Symphypleona) from Brazil, with Notes on the Systematics of the Family. Diversity 2022, 14, 960. [Google Scholar] [CrossRef]
- Greenslade, P.; Weiner, W.M. Taxonomic papers on Collembola in Zootaxa, 2001 to 2020. Zootaxa 2021, 4979, 215–217. [Google Scholar] [CrossRef]
- Cicconardi, F.; Fanciulli, P.P.; Emerson, B.C. Collembola, the biological species concept and the underestimation of global species richness. Mol. Ecol. 2013, 22, 5382–5396. [Google Scholar] [CrossRef] [PubMed]
- Dukes, C.D.; Janssens, F.; Recuero, E.; Caterino, M.S. Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA). Diversity 2022, 14, 847. [Google Scholar] [CrossRef]
- Christiansen, K. Bionomics of Collembola. Annu. Rev. Entomol. 1964, 9, 147–178. [Google Scholar] [CrossRef]
- Petersen, H.; Luxton, M. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 1982, 39, 288–388. [Google Scholar] [CrossRef]
- Pollierer, M.M.; Scheu, S. Driving factors and temporal fluctuation of Collembola communities and reproductive mode across forest types and regions. Ecol. Evol. 2017, 7, 4390–4403. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, K. Convergence and parallelism in cave Entomobryinae. Evolution 1961, 15, 288–301. [Google Scholar] [CrossRef]
- Verhoef, H.A.; Brussaard, L. Decomposition and nitrogen mineralization in natural and agroecosystems: The contribution of soil animals. Biogeochemistry 1990, 11, 175–211. [Google Scholar] [CrossRef]
- Rusek, J. Biodiversity of Collembola and their functional role in the ecosystem. Biodivers. Conserv. 1998, 7, 1207–1219. [Google Scholar] [CrossRef]
- Potapov, A.; Bellini, B.C.; Chown, S.L.; Deharveng, L.; Janssens, F.; Kovác, L.; Kuznetsova, N.; Ponge, J.-F.; Potapov, M.; Querner, P.; et al. Towards a global synthesis of Collembola knowledge—Challenges and potential solutions. Soil Org. 2020, 92, 161–188. [Google Scholar] [CrossRef]
- Forey, E.; Coulibaly, S.F.M.; Chauvat, M. Flowering phenology of a herbaceous species (Poa annua) is regulated by soil Collembola. Soil Biol. Biochem. 2015, 90, 30–33. [Google Scholar] [CrossRef]
- Winck, B.R.; Chauvat, M.; Coulibaly, S.F.M.; Santonja, M.; Saccol de Sá, E.L.; Forey, E. Functional collembolan assemblages induce different plant responses in Lolium perenne. Plant Soil 2020, 452, 347–358. [Google Scholar] [CrossRef]
- Castaño-Meneses, G.; Palacios-Vargas, J.G.; Cutz-Pool, L.Q. Feeding habits of Collembola and their ecological niche. An. Inst. Biol. Ser. Zool. 2004, 75, 135–142. [Google Scholar]
- Souza, P.G.C.; Medeiros, G.S.; Ferreira, R.L.; Souza-Silva, M.; Bellini, B.C. A Highly Troglomorphic New Genus of Sminthuridae (Collembola, Symphypleona) from the Brazilian Semiarid Region. Insects 2022, 13, 650. [Google Scholar] [CrossRef]
- Maaß, S.; Caruso, T.; Rillig, M. Functional role of microarthropods in soil aggregation. Pedobiologia 2015, 58, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.D.D.; Bellini, B.C.; Rigotti, V.M.; Nunes, R.C.; Menezes, L.S.; Winck, B.R. Diversity Loss of Epigeic Collembola after Grassland Conversion into Eucalyptus Forestry in Brazilian Pampa Domain. Diversity 2022, 14, 490. [Google Scholar] [CrossRef]
- Culik, M.P.; Zeppelini Filho, D. Diversity and distribution of Collembola (Arthropoda: Hexapoda) of Brazil. Biodivers. Conserv. 2003, 12, 1119–1143. [Google Scholar] [CrossRef]
- Zeppelini, D.; Queiroz, G.C.; Bellini, B.C. Collembola in Catálogo Taxonômico da Fauna do Brasil. PNUD. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/470 (accessed on 24 January 2023).
- Betsch, J.M. Éléments pour une monographie des Collemboles Symplyplêones (Hexapodes, Aptérygotes). Mém. Mus. Natl. Hist. Nat. Sér. A Zool. 1980, 116, 1–227. [Google Scholar]
- Christiansen, K.; Bellinger, P. The Collembola of North America. North of Rio Grande, a Taxonomy Analysis, 2nd ed.; Grinnell College: Grinnell, IA, USA, 1998; pp. 1–1520. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Families: Neogastruridae and Brachystomellidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1949; pp. 1–341. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Families: Anuridae and Pseudachorutidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1949; pp. 1–122. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Bilobidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1951; pp. 1–97. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Onychiuridae, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1954; pp. 1–219. [Google Scholar]
- Massoud, Z. Monographie des Neanuridae, Collemboles Poduromorphes apiéces buccales modifiées. In Biologie de l’Amerique Australe, 1st ed.; Delamare Deboutteville, C., Rapoport, E.H., Eds.; Éditions du CNRS: Paris, France, 1967; Volume 3, pp. 7–399. [Google Scholar]
- Dunger, W. Synopses on Palaeartic Collembola, Volume 1. Tullbergiinae. Abh. Ber. Naturkundemus. Gorlitz 1994, 68, 1–71. [Google Scholar]
- Cassagnau, P. Collemboles Palaeonurini primitifs d’Afrique et Madagascar. Ann. Soc. Entomol. Fr. 1996, 32, 7–33. [Google Scholar]
- Weiner, W.M. Generic revision of Onychiurinae (Onychiuridae, Collembola) with a cladistic analysis. Ann. Soc. Entomol. Fr. 1996, 32, 163–200. [Google Scholar]
- Najt, J.; Weiner, W.M. Geographic distribution of Brachystomellinae (Collembola, Neanuridae). Pan-Pacific Entomol. 1996, 72, 61–69. [Google Scholar]
- Jordana, R.; Arbea, J.I.; Simón, C.; Luciáñez, M.J. Fauna Iberica Volume 8: Collembola, Poduromorpha, 1st ed.; Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1997; pp. 1–807. [Google Scholar]
- Pomorski, R.J. Onychiurinae of Poland (Collembola: Onychiuridae), 1st ed.; Polish Taxonomical Society: Wrocław, Poland, 1998; pp. 1–210. [Google Scholar]
- Fjellberg, A. Fauna Entomologica Scandinavica—The Collembola of Fennoscandia and Denmark, Part I: Poduromorpha, 1st ed.; Brill: Leiden, The Netherlands, 1998; pp. 1–184. [Google Scholar]
- Thibaud, J.M.; Schulz, H.J.; Assalino, M.M.G. Synopses on Palaeartic Collembola, Volume 4: Hypogastruridae. Abh. Ber. Naturkundemus. Gorlitz 2004, 75, 1–287. [Google Scholar]
- Weiner, W.M.; Najt, J. Species of Brachystomella (Collembola: Brachystomellidae) from the Neotropical region. Eur. J. Entomol. 2001, 98, 387–413. [Google Scholar] [CrossRef]
- Najt, J.; Weiner, W.M.; Grandcolas, P. Phylogeny of the Brachystomellidae (Collembola)—Were the mandibles ancestrally absent and did they reappear in this family? Zool. Scr. 2005, 34, 305–312. [Google Scholar] [CrossRef]
- Paśnik, G.; Weiner, W.M. First phylogenetic analysis of the tribe Oligaphorurini (Collembola: Onychiuridae) inferred from morphological data, with implications for generic classification. Org. Divers. Evol. 2017, 17, 619–631. [Google Scholar] [CrossRef]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Isotomidae, 1st ed.; Polska Akademia Umiejętności, Acta monographica Musei Historiae Naturalis: Kraków, Poland, 1947; pp. 1–488. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Tribe: Orchesellini, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1957; pp. 1–151. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Tribe: Entomobryini, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1957; pp. 1–126. [Google Scholar]
- Potapov, M. Synopses on Palearctic Collembola, Volume 3: Isotomidae. Abh. Ber. Naturkundemus. Gorlitz 2001, 73, 1–603. [Google Scholar]
- Fjellberg, A. Fauna Entomologica Scandinavica—The Collembola of Fennoscandia and Denmark, Part II: Entomobryomorpha and Symphypleona, 1st ed.; Brill: Leiden, The Netherlands, 2007; pp. 1–264. [Google Scholar]
- Jordana, R. Synopses on Palaeartic Collembola, Volume 7/1: Capbryinae & Entomobryini. Soil Org. 2012, 84, 1–390. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Family: Sminthuridae, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1956; pp. 1–287. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects. Families: Neelidae and Dicyrtomidae, 1st ed.; Polska Akademia Nauk, Państwowe Wydawnictwo Naukowe PWN: Kraków, Poland, 1957; pp. 1–113. [Google Scholar]
- Richards, W.R. Generic classification, evolution, and biogeography of the Sminthuridae of the world (Collembola). Mem. Ent. Soc. Can. 1968, 53, 3–54. [Google Scholar] [CrossRef]
- Bonet, F. Monografia de la familia Neelidae (Collembola). Rev. Soc. Mex. Hist. Nat. 1947, 8, 131–192. [Google Scholar]
- Schneider, C. Morphological review of the order Neelipleona (Collembola) through the redescription of the type species of Acanthoneelidus, Neelides and Neelus. Zootaxa 2017, 4308, 1–94. [Google Scholar] [CrossRef]
- Christiansen, K.; Bellinger, P. Insects of Hawaii—A Manual of the Insects of the Hawaiian Islands, including an Enumeration of the Species and Notes on Their Origin, Distribution, Hosts, Parasites, etc. Volume 15—Collembola, 1st ed.; University of Hawaii Press: Honolulu, HI, USA, 1992; pp. 1–445. [Google Scholar]
Figure 1.
(A) General morphology of a springtail, Lepidonella zeppelini Soto-Adames and Bellini, 2015 (Paronellinae, Entomobryidae, Entomobryomorpha). Numbers represent the thoracic and abdominal segments. The first thoracic segment (prothorax) lacks the dorsal plate (pronotum); (B) a male specimen of Tyrannoseira raptora (Zeppelini and Bellini, 2006) with the furca rested next to the ventral body; (C) the same specimen releasing the furca and jumping (tenaculum omitted). Adapted from [8].
Figure 1.
(A) General morphology of a springtail, Lepidonella zeppelini Soto-Adames and Bellini, 2015 (Paronellinae, Entomobryidae, Entomobryomorpha). Numbers represent the thoracic and abdominal segments. The first thoracic segment (prothorax) lacks the dorsal plate (pronotum); (B) a male specimen of Tyrannoseira raptora (Zeppelini and Bellini, 2006) with the furca rested next to the ventral body; (C) the same specimen releasing the furca and jumping (tenaculum omitted). Adapted from [8].

Figure 2.
General habitus of springtails: (A) Paronellinae, Entomobryidae (Entomobryomorpha); (B) Isotominae, Isotomidae (Entomobryomorpha); (C) Hypogastruridae (Poduromorpha), arrow points to the pronotum; (D) Onychiurinae, Onychiuridae (Poduromorpha); (E) Dicyrtomidae (Symphypleona), thorax and abdomen limits are highlighted; (F) male of Sminthurididae (Symphypleona); (G) Neelinae, Neelidae (Neelipleona), thorax and abdomen limits are highlighted; (H) Neelidinae, Neelidae (Neelipleona). Adapted from [9].
Figure 2.
General habitus of springtails: (A) Paronellinae, Entomobryidae (Entomobryomorpha); (B) Isotominae, Isotomidae (Entomobryomorpha); (C) Hypogastruridae (Poduromorpha), arrow points to the pronotum; (D) Onychiurinae, Onychiuridae (Poduromorpha); (E) Dicyrtomidae (Symphypleona), thorax and abdomen limits are highlighted; (F) male of Sminthurididae (Symphypleona); (G) Neelinae, Neelidae (Neelipleona), thorax and abdomen limits are highlighted; (H) Neelidinae, Neelidae (Neelipleona). Adapted from [9].

Figure 3.
Some different phylogenetic hypotheses for the relationships of the Collembola orders found by recent studies (only topologies in which the orders were considered monophyletic are represented). Trees after the mentioned authors: (A) Bellini et al. [9]; (B) Gao et al. [20], Bellini et al. [9]; (C) Schneider et al. [21], Schneider and D’Haese [22]; (D) Leo et al. [23].
Figure 3.
Some different phylogenetic hypotheses for the relationships of the Collembola orders found by recent studies (only topologies in which the orders were considered monophyletic are represented). Trees after the mentioned authors: (A) Bellini et al. [9]; (B) Gao et al. [20], Bellini et al. [9]; (C) Schneider et al. [21], Schneider and D’Haese [22]; (D) Leo et al. [23].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bellini, B.C.; Weiner, W.M.; Winck, B.R. Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue. Diversity 2023, 15, 221. https://doi.org/10.3390/d15020221
AMA Style
Bellini BC, Weiner WM, Winck BR. Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue. Diversity. 2023; 15(2):221. https://doi.org/10.3390/d15020221
Chicago/Turabian StyleBellini, Bruno Cavalcante, Wanda Maria Weiner, and Bruna Raquel Winck. 2023. "Systematics, Ecology and Taxonomy of Collembola: Introduction to the Special Issue" Diversity 15, no. 2: 221. https://doi.org/10.3390/d15020221
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.