

















Ruff Calidris pugnax Scientific name definitions
- LC Least Concern
- Names (63)
- Monotypic
Text last updated March 22, 2017
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Afrikaans | Kemphaan |
| Albanian | Luftëtari |
| Arabic | دريجة شرسة |
| Armenian | Կռվակտցար |
| Asturian | Combatiente |
| Azerbaijani | Döyüşkən cüllüt |
| Basque | Borrokalaria |
| Bulgarian | Бойник |
| Catalan | batallaire |
| Chinese | 流蘇鷸 |
| Chinese (Hong Kong SAR China) | 流蘇鷸 |
| Chinese (SIM) | 流苏鹬 |
| Croatian | pršljivac |
| Czech | jespák bojovný |
| Danish | Brushane |
| Dutch | Kemphaan |
| English | Ruff |
| English (United States) | Ruff |
| Faroese | Kápugrælingur |
| Finnish | suokukko |
| French | Combattant varié |
| French (France) | Combattant varié |
| Galician | Pilro combatente |
| German | Kampfläufer |
| Greek | Μαχητής |
| Gujarati | ટીલિયો |
| Hebrew | לוחם |
| Hungarian | Pajzsoscankó |
| Icelandic | Rúkragi |
| Indonesian | Trinil rumbai |
| Italian | Combattente |
| Japanese | エリマキシギ |
| Korean | 목도리도요 |
| Latvian | Gugatnis |
| Lithuanian | Gaidukas |
| Malayalam | ബഹുവർണ്ണൻ മണലൂതി |
| Marathi | शेरटा |
| Mongolian | Ноцоо элсэг |
| Norwegian | brushane |
| Persian | تلیله شکیل |
| Polish | batalion |
| Portuguese (Angola) | Combatente |
| Portuguese (Brazil) | combatente |
| Portuguese (Portugal) | Combatente |
| Romanian | Bătăuș |
| Russian | Турухтан |
| Serbian | Sprudnik ubojica |
| Slovak | bojovník bahenný |
| Slovenian | Togotnik |
| Spanish | Combatiente |
| Spanish (Costa Rica) | Combatiente |
| Spanish (Cuba) | Combatiente |
| Spanish (Ecuador) | Combatiente |
| Spanish (Mexico) | Playero Combatiente |
| Spanish (Panama) | Combatiente |
| Spanish (Peru) | Playero Combatiente |
| Spanish (Puerto Rico) | Combatiente |
| Spanish (Spain) | Combatiente |
| Spanish (Venezuela) | Rufo |
| Swedish | brushane |
| Thai | นกรัฟ |
| Turkish | Döğüşkenkuş |
| Ukrainian | Брижач |
Calidris pugnax (Linnaeus, 1758)
Definitions
- CALIDRIS
- calidris
- pugnax
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
Male 26–32 cm, 130–254 g, wingspan 54–58 cm; female 20–25 cm, 70–170 g, wingspan 48–52 cm. Extreme sexual dimorphism. Male has head-tufts and ruff variably coloured buff, chestnut, dark purple, black or white , often barred or flecked; white coloration of tufts and ruff associated with satellite behavioural role; mantle and scapulars vary from black to brown, buff, chestnut, ochre or white; yellow to brown facial warts; underparts usually dark, lower belly and undertail white; bill brown to dull orange and legs yellow-green to dark orange. There also appear to exist, however, some permanently female-like males (1). Male nuptial ornament diversity extensive and ruff , head-tufts and facial wattles differ in coloration and pattern individually . An analysis of 1814 males caught in their spring staging areas in Friesland, Netherlands, identified 801 different variants . Individual feathers were found to be plain (white, black or one of a range of hues), or patterned with black and just one hue. The colour of a male’s ruff plus head-tufts consisted of black and/or white and/or only one other hue. Nevertheless, except wattle colour, characteristics did not combine randomly. Some combinations , such as a white ruff with white hea= tufts, occurred much more frequently than expected by chance. Other combinations, such as a white ruff with black head-tufts , a black ruff with white hea- tufts , and a regular ruff with an irregular pattern, were rare. Mostly there was conformity between ruff and head-tufts: they were identical or had reversed primary and secondary colours (2). Female considerably smaller than male , with overall colour rather variable; lacks specialized head plumage of male; dark upperparts with buff to rufous fringes and black blotches on breast and flanks; bill black, legs pinkish orange to green or grey. Non-breeding adult like breeding female, but paler grey-brown above with whiter lower face and dusky buff on breast; bill dark, legs duller; some males retain much white on head and neck. Juvenile has buff-fringed dark brown upperparts and rufous-buff foreneck, breast and belly; face mainly buff with pale throat; bill and legs as in breeding female.
Systematics History
Subspecies
Distribution
NW & N Europe E through Siberia to Chukotskiy Peninsula and Sea of Okhotsk. Winters from W Europe, Mediterranean and sub-Saharan Africa through Middle East to Indian Subcontinent and SE Asia, also Greater Sundas and Philippines.
Habitat
Breeds in Arctic and subarctic, and S to temperate zones in Europe; requires habitat with adjacent feeding, lekking and nesting areas; coastal tundra to forest tundra, near small lakes, in marshes and deltas with shallow-water margins, variably covered with vegetation, and dry mounds and slopes with some low scrub for leks; in W Europe, damp to swampy meadows, often with shallow pools or ditches, and patches of birch, willow scrub and short grass or bare ground. Outside breeding season, prefers muddy margins of lakes, pools, ponds, rivers, marshes and flooded areas , including brackish, saline or alkaline waters; in India, large numbers on tidal mudflats and lagoons, but not commonly on seashore elsewhere; W African wintering sites include flooded or dry plains, marshes and grass, wheat or rice fields, not always close to water; observed picking at spilled grain on road. During migration commoner along shores and in freshly mown or short-grazed grasslands in W Europe. Uses night-time roosts on shallow water along lake edges.
Movement
Migratory. Chief winter quarters in Africa; huge concentrations in Sahel zone, e.g. Senegal Delta, Niger inundation zone in Mali, Sokoto region in Nigeria, Yobe Delta in Niger and Nigeria, Nile system in Sudan. In autumn, moves on broad front SW across Europe, following route further E in spring; Scandinavian breeders probably virtually all stage in Wadden Sea region from late Apr to mid May; W movement of E breeders in autumn towards Europe; E & S African winterers come from Siberia, migrating via Caspian and Black Seas; smaller numbers fly across easternmost Asia. Autumn migration starts late Jun to early Jul (males), mid Jul (females) and late Jul to Aug (juveniles), crossing temperate Europe mainly late Jul to mid Sept, and S Asia Sept; at least in Britain juvenile females precede juvenile males; important staging and moulting areas around North Sea, W Germany and N Caspian Sea; arrives Senegal from mid Jul (earliest males) and tropics mainly late Aug to Sept. Females winter farther S than males and juveniles move S gradually. Spring migration starts Mar, but mainly Mar to mid May, males starting a month earlier than females; some non-breeders remain in winter quarters; breeding grounds reoccupied from mid Apr in W Europe, but progressively later to N & E, mid May to Jun in Siberia. Migrates in large groups, of 100s or 1000s.
Diet and Foraging
During breeding season, mainly terrestrial and aquatic insects, including adults and larvae, particularly of beetles and dipteran flies; outside breeding season diet more varied, including caddisflies, water-bugs, mayflies and grasshoppers, small crustaceans, spiders, small molluscs, annelid worms, frogs and small fish; birds wintering in Sahel zone often feed on rice seeds and other cereals , sedges, grasses and aquatic plants. Probes in mud or soil and picks prey from surface or plants; sometimes follows ploughs or wades in shallow water. Nocturnal and diurnal feeder.
Sounds and Vocal Behavior
Hardly vocal, even on breeding grounds, where during extensive display on lek remains silent. Voice basically limited to short, guttural, subdued notes.
Breeding
Lays from early May in NW Europe, mid May in Sweden and early Jun in Finnmark. Polygynous, with no true pair-bond; mating primarily occurs at traditional leks where males gather to display and females visit. Three types of males appear on arenas: (1) residents, possessing small territories (‘courts’); (2) marginals, without territory; and (3) satellites, occurring on periphery and mating opportunistically; satellites generally have white tufts and ruffs. It has proved impossible to correlate male plumage characteristics with female interest or mating success . Has been suggested that plumage diversity is useful to signal status (satellite or independent male); satellite males benefit from signalling their mating strategy and their white plumage enables admission to mating sites and, in combination with particular postures, restrains territory holders from attacking them. Plumage diversity may also serve to signal individual identity, in a species that does not sing. Being identifiable is beneficial for territorial males that have settled on a lek ; their neighbours have tested their fighting abilities and will avoid their territory. This minimizes violence to ritual attacks or very short fights. In general, however, the evolutionary drivers of extreme diversification in males remain largely unknown (2). Males of an additional female-like morph described in 2006 from Netherlands; intermediate-sized, permanently female-plumaged individuals represent an odd example of ‘sexual mimicry’, a strategy used to gain access to females without inducing aggression from other males (1). Many females may copulate during migration. No site fidelity and natal philopatry. Nests solitary or semi-colonial, in densities of 0·1–2·5 nests/ha, occasionally only several metres apart and sometimes near lek; non-territorial. Nest concealed in marsh plants or meadow grass; shallow scrape lined with grass, leaves and stems. Clutch four eggs, sometimes three, laid at 24–36 hour intervals; incubation 20–23 days, starting with last egg; incubation and brood rearing by female only; chick ochre-orange above, with black and brown streaks and lines of ‘powder-puffs’, of which two along crown, underparts entirely buff; fledging 25–28 days; female leaves chicks at or some days before fledging. Hatching success 93% (only one study). Age of first breeding two years, probably later in males; annual mortality 48%; oldest ringed bird ten years, 11 months.
Conservation Status
Not globally threatened (Least Concern). Total population possibly more than 2,000,000 birds, of which > 1,000,000 winter in W Africa, up to 1,000,000 in E Africa and SW Asia; up to 1,000,000 birds counted within a 25 km radius of Senegal Delta in 1972, 180,000–200,000 in 1991 and 1992; fewer than 10,000 birds in Atlantic Europe and Mediterranean; c. 100,000 in SC Asia, with 113,000 in India in 1992. In Norway and Sweden, 60,000–95,000 breeding ‘pairs’ and 30,000–50,000 in Finland, where earlier estimated at 196,000 in 1973–1976 and 150,000 in 1986. Numbers declining at W African wintering sites. W breeding range has contracted considerably during last 200 years, especially in S, as result of drainage of wetlands; in Netherlands decline of 90% since 1950s. In Norway and Finland, has expanded range S since 1940s. In recent years breeding range and numbers have increased in E Russia; small British population has increased since late 1960s.
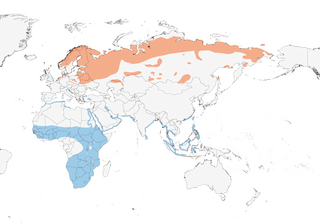
- Year-round
- Migration
- Breeding
- Non-Breeding



















