










Northern Lapwing Vanellus vanellus Scientific name definitions
- NT Near Threatened
- Names (49)
- Monotypic
Text last updated April 13, 2017
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Gicvilja |
| Arabic | قطقاط شمالي |
| Armenian | Սովորական կիվիվ |
| Asturian | Pegueta europea |
| Azerbaijani | Adi çökükburun |
| Basque | Hegabera |
| Bulgarian | Обикновена калугерица |
| Catalan | fredeluga europea |
| Chinese | 小辮鴴 |
| Chinese (Hong Kong SAR China) | 鳳頭麥雞 |
| Chinese (SIM) | 凤头麦鸡 |
| Croatian | vivak |
| Czech | čejka chocholatá |
| Danish | Vibe |
| Dutch | Kievit |
| English | Northern Lapwing |
| English (United States) | Northern Lapwing |
| Faroese | Vípa |
| Finnish | töyhtöhyyppä |
| French | Vanneau huppé |
| French (France) | Vanneau huppé |
| Galician | Avefría choromica |
| German | Kiebitz |
| Greek | (Ευρωπαϊκή) Καλημάνα |
| Hebrew | קיווית מצויצת |
| Hungarian | Bíbic |
| Icelandic | Vepja |
| Italian | Pavoncella |
| Japanese | タゲリ |
| Korean | 댕기물떼새 |
| Latvian | Ķīvīte |
| Lithuanian | Paprastoji pempė |
| Mongolian | Умардын хавтгаалж |
| Norwegian | vipe |
| Persian | خروس کولی معمولی |
| Polish | czajka |
| Portuguese (Portugal) | Abibe |
| Romanian | Nagâț |
| Russian | Чибис |
| Serbian | Vivak |
| Slovak | cíbik chochlatý |
| Slovenian | Priba |
| Spanish | Avefría Europea |
| Spanish (Puerto Rico) | Avefría Europea |
| Spanish (Spain) | Avefría europea |
| Swedish | tofsvipa |
| Thai | นกกระแตหงอน |
| Turkish | Kızkuşu |
| Ukrainian | Чайка чубата |
Vanellus vanellus (Linnaeus, 1758)
Definitions
- VANELLUS
- vanellus
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
28–31 cm; 128–330 g; wingspan 82–87 cm. Metallic glossy green upperparts , blackish crest and bronze scapulars; very broad wings , especially in breeding males. Female has less distinctive head pattern and white flecks on throat. Non-breeding adult has buff face, short crest, and white chin and throat; upperwing-coverts and scapulars have buff fringes. Juvenile similar to non-breeding adult, but has more extensive buff feather fringes , and narrower and browner breastband.
Systematics History
Subspecies
Distribution
Europe, Turkey and NW Iran through W Russia and Kazakhstan to S & E Siberia, Mongolia and N China. Winters from W Europe, E Atlantic islands and N Africa through Mediterranean, Middle East and Iran across N India to SE China, Korea and S Japan.
Habitat
Breeds in variety of wide open habitats with short vegetation or bare ground, including various wetlands , heaths, moors, arable and cereal fields, meadows and hayfields, often with other wading birds such as Black-tailed Godwits (Limosa limosa), Common Snipes (Gallinago gallinago), Dunlins (Calidris alpina), Ruffs (C. pugnax) and Common Redshanks (Tringa totanus). In Europe mainly a farmland breeder , but recently recorded nesting on a forestry clear-cut in Sweden (1). Breeds to c. 1000 m in Europe, but mainly below c. 800 m; in Indian Subcontinent in non-breeding season mainly below 1300 m (2) (though recorded to 2800 m in Bhutan) (3) and observed up to 3350 m in Sichuan, SW China (4). Outside breeding season also appears on harvested stubble and ploughed fields, frequently in company of Pluvialis apricaria, and is increasingly using arable fields in winter as opposed to pastureland, at least in parts of NW Europe (5). Roosting flocks prefer spacious, old pastures and sometimes appear on mudflats, as well as (recently) on large open roofspaces (6).
Movement
Migratory and resident. Post-breeding gathers in flocks . Breeding areas vacated as early as beginning of summer, movements starting already in early Jun. Movements during summer mainly westward. British and southernmost breeders partially sedentary; many British birds winter in Ireland. Migration on broad front towards SW takes place in Sept–Nov, and starts with cold spells, with Finnish breeders moving S to Italy or N Africa. Wintering area bordered by 3°C isotherm, but exact S & W distribution depends on weather, although regularly reaches most of Macaronesia (even the Azores), except the Cape Verdes, where very rare (7). In SE Asia, scarce as far S as Taiwan at this season (8), but also reaches Myanmar, Thailand and Indochina (9), while numbers reaching N Indian Subcontinent also strongly influenced by relative weather conditions (2), although species appears to be occurring more regularly in Bangladesh in recent years (10). Spring migration at peak in early Mar, but can start as early as Jan, with evidence for earlier arrival in recent decades in parts of range (11, 12) and an advancing breeding schedule (compared to dates in 1960s) as a result (13); males precede females and breeding grounds typically reoccupied by Apr. Vagrants have reached as far afield as Spitsbergen, Bear I, Jan Mayen, Senegambia (14), Kenya (15), the Philippines (three records, Nov–Jan) (16), Borneo (three records, Nov–Jan) (17), Alaska (one autumn record), E North America (almost annual late Oct–Mar in Newfoundland, sometimes in large numbers, but very rare elsewhere in this region W to Ohio and S to Florida and Georgia, mainly during same period though also recorded in summer) (18, 19), Bermuda, and the Caribbean (Bahamas, Puerto Rico, Martinique and Barbados) (20, 21).
Diet and Foraging
Feeds on invertebrate prey, primarily earthworms and insects, including larvae and adults of beetles, ants, Diptera flies, moths and crickets; also spiders and snails. On arable fields, adults and chicks take mainly cranefly larvae and earthworms, but evidently not selectively. Study in UK found that proportion of aquatic invertebrates in diet of both species increases as breeding season progresses (22). Winter study in E England found that numerically, arthropods (mostly Carabids and millipedes) were main diurnal prey types but, by biomass, small earthworms were major prey items and that nocturnal intake rates were up to 50% higher due to greater reliance on catching large earthworms at night; it also reported that diurnal intake rates were highest during mild weather and on grass and sugar beet stubble, but were lowest on cereal crops, despite this habitat being that most consistently occupied (23). Locates prey both visually and aurally, mostly as bird walks or runs, occasionally pausing to probe ground. Rather frequently feeds by foot-trembling. Diurnal and nocturnal; sometimes even primarily nocturnal on bright, moonlit nights, but flocks are more dispersed when feeding at night (24). Kleptoparasitised by Black-headed Gulls (Larus ridibundus) (22).
Sounds and Vocal Behavior
Very vocal during breeding season when male’s territorial song given (see also Breeding) during display flight is a highly distinctive, but complex and twangy “ayhrre-willuch-o-weep”, in which the first note is slurred (and easily missed), and contact/alarm calls are equally recognizable, a “pee-wi” or “peeawee”, which gave rise to its older English name, Peewit. In non-breeding season, most regularly heard are a loud, shrill “cheew” or a more plaintive-sounding “cheew-ip” or “wee-ip”, sometimes rising and falling in pitch (a similar call is also given by adults in flight with nest or young in alarm).
Breeding
Lays mid/late Mar to early Jun overall, with start becoming progressively later in N & E of range. Mostly seasonally monogamous, but sometimes polygamous; in SW Norway, 23–41% of males were polygynous, having two, sometimes three and exceptionally four (25) mates, whereas 50–77% were monogamous and 0–21% remained unmated (26), while in NE England 30% of males (especially older than second-year) had more than one mate, while one female (of 64 studied) was polyandrous (27). Territorial during incubation; solitary breeder or loosely colonial, with mean densities 0·6–1·8 pairs/ha, maximum 9 pairs/ha. High degree of site-faithfulness in males, and high degree of natal philopatry, usually nesting within 60 km of hatching site. Males perform dazzling aerial song flight display (see Family Text), during which wingtips produce droning sound (“wup-wup-wup”) and includes a rolling section, as well as dramatic dives and switchbacks (28). Females may sample many males and territories in the course of a short time span (29). Nest a shallow scrape, lined with some (occasionally substantial) vegetation, sited in short grassy vegetation, in wet areas on slight eminence (30). Single-brooded, but will lay replacement clutch if first lost (30) and double-brooding occasionally recorded, with male taking exclusive responsibility for first brood after young hatch and female for the second (31, 32). Clutch four eggs , occasionally 2–3 or five, laid on consecutive or alternate days, colour creamy buff or stone, rarely bluish or reddish, marked with black, mean size 47·1 mm × 33·7 mm (30) (range 42·7–50·9 mm × 31·1––35·8 mm in one study) (33); clutch completed in five days; incubation 21–30 days (34), by both sexes (mainly female, especially those partnered with bigamous males) (35, 30, 36), starting once last egg laid; downy chick pale brown or cinnamon, with black streaks and blotches and white nape (mean mass 17·5 g) (37); chicks tended by both parents, although brooding primarily by female; fledging 33–40 days. Some evidence of intraspecific egg-dumping (38) and one instance of apparent interspecific parasitism, by Eurasian Oystercatchers (Haematopus ostralegus) (39), while a study in Poland reported that lapwings will practice egg-dumping on Black-tailed Godwits (Limosa limosa) and Common Redshanks (Tringa totanus) (40). Agricultural activities may cause 36% of broods to fail, plus another 32% partial losses of clutches; wet and cold weather can have deleterious effects during chick phase (41); other causes of failure are predation, especially by crows, trampling by cattle and flooding, while presence livestock can have additional knock-on effects, e.g. disturbing incubating birds, permitting predators additional opportunities to seize eggs (42). Study in Netherlands identified c. 15 species of predators of eggs and chicks, of which red foxes (Vulpes vulpes), Grey Herons (Ardea cinerea) and stoats (Mustela erminea) were among the most frequent (37). In optimal habitat, hatching success can be in excess of 70% (43) and overall breeding success estimated at c. 61% in Russian study (33), whereas in contrast hatching success just 21% in SE Sweden (44). Highest mortality of hatchlings generally during first ten days, but can remain high throughout fledging period (45). Fledging success varies with year and habitat, but averages 0·2–2·8 fledglings per pair, with multi-year Dutch study reporting maximum of 23% success rate (45). Study in NE England found that average productivity of males increases by c. 40% between one and two years of age and by another 10% between two and three years old, reflecting a 40% increase in proportion of males that breed between one and two years old and an increase in mating success with age coupled with higher breeding success for polygynous males (46), although in SW Norway researchers found that secondary females often fared poorly (47). Age of first breeding mostly one year; annual mortality of adults c. 25–30%, first-year birds c. 35–40%. Oldest ringed bird 18 years, 11 months.
Conservation Status
Not globally threatened. Currently considered Near Threatened. Global population thought to number up to 11,000,000 individuals (2006 data) (48). Estimated 1,590,000–2,580,000 pairs in Europe, of which 500,000–850,000 in Russia, 126,472–286,669 in Latvia, 131,655–229,856 in Netherlands, c. 63,000 in UK (in 1998, a 49% decline since 1987) (49), 100,000–160,000 in Belarus, 92,000–120,000 in Finland, 90,000–120,000 in Poland, 65,000–130,000 in Romania, 63,000–100,000 in Germany (2005–2009 data) (50), and > 20,000 pairs each in Ukraine, Sweden, Hungary and Estonia (51). Very rare breeder in Faeroes and Iceland, while populations at S edge of breeding range (Morocco, Israel) have noticeably declined or become extinct, respectively. More than 25,000 birds winter in SW Asia, c. 20,000 birds in Iran, 10,000–25,000 birds in SC Asia; more than 25,000 birds in E & SE Asia (including c. 7500 in lower R Yangtze floodplain) (52), and this population is increasing. In winter (Nov), 1,000,000–1,500,000 birds present in Netherlands and 100,000–200,000 in Denmark; in Jan, 314,000 in Britain (Jan counts in 1990s) (53), where population has shifted eastwards at this season since 1980s (54), although 70,000 gathered on Somerset Levels in SW England in winter 2010/11 (55). Breeding area expanding northwards throughout range, mainly following extensive, man-made water meadows. However, European population decreased in 19th and early 20th centuries, probably as result of intensification of land use, drainage and egg-collecting, and same now happening in many E parts of range; due to habitat shift to agricultural land, numbers thereafter increased, until recent decades, when population of W Europe in decline again, apparently caused by intensifying and changing agricultural practices, resulting in reduced breeding productivity (56, 57, 58); on arable fields, clutch destruction during cultivation and poor chick survival can result in insufficient productivity to maintain a stable population, while loss of spring tillage may also be a contributory factor (59). Even on organic farms, where densities may be higher, nesting success can be still be comparatively low (60). Retaining high water level tables throughout the breeding season appears to be an important condition of nesting success (61, 41) and is unlikely to be achieved in many areas without highly proactive management, while measures to prevent ground-dwelling predators accessing chick-rearing areas have also been tested with some success (62). Marked decreases in recent decades in SW Norway (63), S Sweden, and in large parts of Britain (e.g., 38% decline in lowlands of England and Wales between 1982 and 2002) (64, 65), Netherlands, Germany (e.g. in Schleswig-Holstein) (66) and Czech Republic (67, 68). European population has fallen by 55% since 1980 (69), and estimated to be still decreasing by 30–49% in 27 years (three generations) (51), despite partial success of agri-environment schemes, funded by the European Union, designed to assist this and other bird species, in part because implementation of such projects has been too localized and also because some species possess complex ecological requirements (70, 71, 72). A similar ‘large decline’ was noted in African–Eurasian flyway population during the period 1988–2012 (73). Given widespread and sustained population declines, since 2015 this species is considered Vulnerable in Europe (51), and globally Near Threatened. A European management plan was published in 2009 (74).
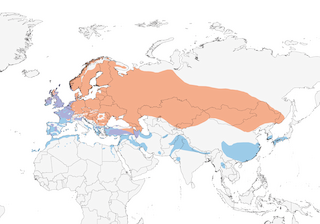
- Year-round
- Migration
- Breeding
- Non-Breeding


eBird and Macaulay Library

eBird Status and Trends

















