













Long-tailed Tit Aegithalos caudatus Scientific name definitions
Text last updated February 5, 2013
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Trishtili bishtgjatë |
| Armenian | Երկարագի երաշտահավ |
| Asturian | Raullargu europñu |
| Azerbaijani | Uzunquyruq arıquşu |
| Basque | Buztanluzea |
| Bulgarian | Дългоопашат синигер |
| Catalan | mallerenga cuallarga eurasiàtica |
| Chinese (Hong Kong SAR China) | 銀喉長尾山雀 |
| Chinese (SIM) | 北长尾山雀 |
| Croatian | dugorepa sjenica |
| Czech | mlynařík dlouhoocasý |
| Danish | Halemejse |
| Dutch | Staartmees |
| English | Long-tailed Tit |
| English (United States) | Long-tailed Tit |
| Finnish | pyrstötiainen |
| French | Orite à longue queue |
| French (France) | Orite à longue queue |
| Galician | Subeliña común |
| German | Schwanzmeise |
| Greek | Αιγίθαλος |
| Hebrew | ירגזי זנבתן |
| Hungarian | Őszapó |
| Icelandic | Skottmeisa |
| Italian | Codibugnolo |
| Japanese | エナガ |
| Korean | 오목눈이 |
| Latvian | Garastīte |
| Lithuanian | Ilgauodegė zylė |
| Mongolian | Сүүлт хөх бух |
| Norwegian | stjertmeis |
| Persian | چرخریسک دم دراز |
| Polish | raniuszek |
| Portuguese (Portugal) | Chapim-rabilongo |
| Romanian | Pițigoi codat |
| Russian | Ополовник |
| Serbian | Dugorepa senica |
| Slovak | mlynárka dlhochvostá |
| Slovenian | Dolgorepka |
| Spanish | Mito Común |
| Spanish (Spain) | Mito común |
| Swedish | stjärtmes |
| Turkish | Uzun Kuyruklu Baştankara |
| Ukrainian | Синиця довгохвоста |
Aegithalos caudatus (Linnaeus, 1758)
Definitions
- AEGITHALOS
- caudata / caudatus
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
13–16 cm, of which tail 6–10 cm; 6.2–10.4 g (europaeus). A tiny, round-bodied passerine with short stubby bill and very long, narrow tail; plumage mainly black and white, with variable amounts of gray and pink.Nominate subspecies has head, throat and most of underparts snow-white , flanks to vent clean pink; upper mantle black, lower mantle and back mixed black and dull pinkish-white to whitish, scapulars and rump pale pink-tinged white with black and white mottling; uppertail-coverts and tail black, outer three rectrices extensively white (especially on outer web); upperwing-coverts and alula black, inner greater coverts broadly tipped and fringed white; flight-feathers dark brown, tertials and secondaries broadly fringed whitish; iris dark hazel, eyelid yellow to red (yellow commonest, but color probably also reflecting mood); bill black; legs blackish-brown. Sexes alike. Juvenile is darker overall than adult, rear lores to cheek and ear-coverts dusky (accentuating pale eyering), only center of forehead and crown whitish, mantle and back brown, pinkish tone confined to distal half of scapulars, tail shorter and blackish-brown, white in outer tail reduced and sullied brownish, inner greater coverts more narrowly fringed and tipped buffish-white, flight-feathers paler and browner, underparts dirty white, belly with buff tinge. Subspecies exhibit marked variation, mostly in plumage pattern and tones, “europaeus group” and “trivirgatus group” slightly duller than nominate and with broad blackish lateral crownstripe, but great individual variation, “alpinus group” also very variable, head pattern as “europaeus group” (although sides of head and neck streaked darker) but upperparts mainly gray with little or no pink, blackish confined to band across upper mantle, often a blackish spot on throat, and has wing and especially tail shorter: rosaceus is smaller than nominate, has forehead to nape white, irregularly spotted dark brown, band from just in front of eye (occasionally from bill) to side of nape dull black, ear-coverts and side of neck finely streaked dark brown, pale areas above darker dull pink, less white in wing (edgings narrower), underparts off-white, flanks dull vinaceous pink, sometimes faint necklace of darker streaks across upper breast; aremoricus has center of crown and underparts whiter than previous, lateral crownstripes broader and blacker; <em>taiti</em> is small and dark, with lateral crownstripes even broader and blacker, upperparts (especially rump) blacker, with less pink; macedonicus resembles previous, but gorget better marked, and lateral crownstripe extends farther forward, often to bill; europaeus typically resembles rosaceus but is brighter and paler, with lateral crownstripe narrower, crown, cheek, ear-coverts and underparts clearer white, but very variable, lateral crownstripe sometimes broken or almost absent (thus resembling nominate, but always at least some indication of dark stripe) or very broad (leaving only small white patch in center of crown); tauricus resembles last, but lateral crownstripe blacker and better defined, mantle blacker with reduced (and paler) pink, scapulars often grayish; <em>trivirgatus</em> also is similar to europaeus, but white areas purer white, black on upperparts restricted to upper mantle, lower mantle paler and with greater admixture of brighter pink; kiusiuensis is like previous but darker, with gorget less distinct or lacking; <em>magnus</em> is close to nominate, but with blackish lateral crownstripe and usually a well-marked gorget of blackish spots; <em>irbii</em> has narrow black band across upper mantle, rest of mantle and scapulars gray, scapulars sometimes tinged pink; italiae is similar to previous, but broader black band on upper mantle , slightly darker slate-gray lower mantle , and with scapulars more broadly tipped pink; siculus also is similar, but lateral crownstripe duller and browner, white center of crown washed buff, dark band on upper mantle reduced and more brownish-black, rest of upperparts paler and grayer, virtually lacking pink, may show vague sooty-black spot on lower throat; tephronotus resembles previous, but lateral crownstripe black, dark upper-mantle band lacking or reduced to a small brown spot, underparts buffish with well-defined blackish throat spot; major is similar, but lateral crownstripe blackish-brown, center of crown whiter (not buffish) with a few small brown streaks, band on upper mantle brown to black and varying in width, rump bright pink, breast whiter, lower belly and flanks brighter pink, dark spot on lower throat faint or absent, tail relatively long; alpinus is like tephronotus, but center of crown brown-streaked buffish-white, upperparts darker, upper mantle gray with some admixed black, lower throat usually sooty, underparts washed buffy brown (pink flanks contrasting little); passekii resembles last, but overall rather paler, especially below, with only faint brown wash on belly, center of crown white (not buff), and lacks gorget (although has dark throat spot).
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Often treated as conspecific with A. glaucogularis, but plumage distinctive, and no reports of regular intergradation or hybridization with latter species in NE China (apparent “intergrades” recorded as migrants in coastal Hebei, but origin and status of these unknown). Marked geographical variation, although the two main vocalizations almost identical throughout range (1). Races fall into four groups: “caudatus group” in N Europe and Asia, “europaeus group” in S & W Europe, “alpinus group” in Mediterranean Europe and SW Asia, and “trivirgatus group” in S Korea and Japan (“europaeus group” and “trivirgatus group” are morphologically similar and usually combined, but marked geographical separation strongly suggests different origins). Where groups meet there are extensive areas occupied by variable populations of apparent hybrids; these areas of hybridization considered zones of secondary intergradation (where populations have met relatively recently). Nominate race intergrades with europaeus in Denmark, E Germany, S Poland, S Ukraine and N Romania, and with magnus in Korea; aremoricus intergrades with taiti in WC France; taiti intergrades with irbii in C Spain and Portugal; and europeaus intergrades with aremoricus in W France, with taiti in SW France, with italiae on S slopes of Alps, and with macedonicus in S of former Yugoslavia and S Bulgaria. Birds of this species breeding in Iraq and Syria of uncertain racial identity, provisionally included in tephronotus. Other proposed races are sibiricus (C Siberia), kamtschaticus (Kamchatka, in E Russia) and japonicus (N Japan), all poorly differentiated and thus synonymized with nominate; and pallidolumbo (Shikoku, in S Japan) and tarihoae (Jeju I, off S Korea), both merged with trivirgatus. Seventeen subspecies recognized.Subspecies
Long-tailed Tit (caudatus) Aegithalos caudatus caudatus Scientific name definitions

Distribution
Fennoscandia (north to c. 70°N in Norway, locally to 66°N in Finland) and northeastern Europe (Poland) east in broad band through Siberia to eastern Russia (Kamchatka), Sakhalin, southern Kuril Islands and northern Japan (Hokkaido, Rebun-to and Rishiri-to), extending south to northern Kazakhstan, northern Mongolia, northeastern China and N Korea (south to 40–41°N); non-breeding also south to eastern China (northeastern Hebei, Beijing) and central Japan (south-central Honshu).
Aegithalos caudatus caudatus (Linnaeus, 1758)
Definitions
- AEGITHALOS
- caudata / caudatus
The Key to Scientific Names
Legend Overview
Long-tailed Tit (europaeus Group) Aegithalos caudatus [europaeus Group]


Distribution
Aegithalos caudatus rosaceus Mathews, 1938
Definitions
- AEGITHALOS
- caudata / caudatus
- rosaceus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus aremoricus Whistler, 1929
Definitions
- AEGITHALOS
- caudata / caudatus
- aremoricus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus taiti Ingram, 1913
Definitions
- AEGITHALOS
- caudata / caudatus
- taiti
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus europaeus (Hermann, 1804)
Definitions
- AEGITHALOS
- caudata / caudatus
- europae / europaea / europaeus / europeus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus macedonicus (Salvadori & Dresser, 1892)
Definitions
- AEGITHALOS
- caudata / caudatus
- macedonica / macedonicus
The Key to Scientific Names
Legend Overview
Aegithalos caudatus tauricus (Menzbier, 1903)
Definitions
- AEGITHALOS
- caudata / caudatus
- taurica / tauricus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus magnus (Clark, 1907)
Definitions
- AEGITHALOS
- caudata / caudatus
- magnum / magnus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus trivirgatus (Temminck & Schlegel, 1848)
Definitions
- AEGITHALOS
- caudata / caudatus
- trivirgata / trivirgatus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus kiusiuensis Kuroda, 1923
Definitions
- AEGITHALOS
- caudata / caudatus
- kiusiuensis
The Key to Scientific Names
Legend Overview
Long-tailed Tit (alpinus Group) Aegithalos caudatus [alpinus Group]



Distribution
Aegithalos caudatus irbii (Sharpe & Dresser, 1871)
Definitions
- AEGITHALOS
- caudata / caudatus
- irbii
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus italiae (Jourdain, 1910)
Definitions
- AEGITHALOS
- caudata / caudatus
- italiae
The Key to Scientific Names
Legend Overview
Aegithalos caudatus siculus (Whitaker, 1901)
Definitions
- AEGITHALOS
- caudata / caudatus
- sicula / siculus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus tephronotus (Günther, 1865)
Definitions
- AEGITHALOS
- caudata / caudatus
- tephronota / tephronotum / tephronotus
- Tephronotus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus major (Radde, 1884)
Definitions
- AEGITHALOS
- caudata / caudatus
- major
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus alpinus (Hablizl, 1783)
Definitions
- AEGITHALOS
- caudata / caudatus
- alpina / alpinum / alpinus
The Key to Scientific Names
Legend Overview
Distribution
Aegithalos caudatus passekii (Zarudny, 1904)
Definitions
- AEGITHALOS
- caudata / caudatus
- passekii
The Key to Scientific Names
Legend Overview
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Deciduous and mixed woodland with well-developed shrub layer, especially willows (Salix), favouring edge habitats. Also riverine woodland and, perhaps especially in W Europe, scrub, heathland with scattered trees, bushes and hedges in farmland, and well-wooded suburban parks, cemeteries and gardens . In Mediterranean region also in maquis and open pine (Pinus) forest. Otherwise, avoids pure stands of conifers in W of range, but noted in open forests of spruce (Picea), fir (Abies), larch (Larix) and Siberian stone pine (Pinus sibirica) in C Siberia (probably especially forest edge or riverside areas where an admixture of broadleaf trees). In Europe usually below 1000 m, although found at up to 1200 m in Hungary and very locally to 1800 m (rarely, to 2120 m) in the Alps, but more montane in SE, breeding to 2000 m in Greece (though commonest at 200–1000 m), 2000 m (mainly 100–1300 m) in Turkey and 1500 m in Caucasus; to at least 1830 m in Iran; in Japan mainly at 500–1600 m, wandering to lower levels in winter; primarily montane also in China, where noted at 780–1100 m (extremes 400–1800 m) in the Changbai Shan (Jilin).
Movement
Resident in temperate and Mediterranean regions, but partially migratory farther N. Birds from N Siberia undertake regular migration late Aug, returning in May, with some dispersal to N in autumn (when flocks of hundreds of individuals recorded in C Siberia), although bulk of movement in S direction; numbers in S Siberia swelled by immigrants in winter, when species may be numerous as far S as Zaysan depression, in NE Kazakhstan (migrants recorded also in R Ili valley). Some remain in N of range (or even farther N than breeding range). In NE China, an irregular migrant and winter visitor at Beidaihe (coastal Hebei), Sept–Feb, occurring in larger numbers during “irruptions”. White-headed birds (nominate race) occur in winter S of breeding range in China, Japan and irregularly S to extreme S Korea (Hong Kong records presumed to involve escapes from captivity). N populations also make irregular, sometimes large-scale irruptions in W, when large numbers may reach Finland, Baltic region and E Europe, and much smaller numbers appear in W Europe, such movements reaching peak in mid-Oct and at least some individuals returning to natal area in spring; birds from N & E sometimes penetrate farther W (e.g. flock of 200 white-headed birds in Netherlands in Dec 1971), but crossing of water avoided, thus nominate race very rarely recorded in Britain, and within British Is resident rosaceus only an irregular autumn wanderer to Is of Scilly, I of May, Outer Hebrides and Orkney, and very rarely to Shetland. Vagrant in Morocco (Apr 1967). Populations breeding in mountainous areas presumably undertake some altitudinal movement, spreading to adjacent foothills and plains in winter (e.g. in Hebei, in NE China).
Diet and Foraging
Food throughout year largely invertebrates ; occasionally also seeds, buds and even olives (Olea) and peanuts taken, especially in autumn and winter; recorded as drinking sap. Forages mostly in shrub layer and canopy, only occasionally on ground, delicately picking items from surface of twigs, leaves and buds. Can hover, and sometimes flycatches. When dealing with an insect too big to be swallowed immediately, will hang from a twig by one foot and hold the item in other foot in order to peck at it. Tame and very gregarious, for most of year forming stable flocks of c. 3–30 individuals based around parents and offspring of previous breeding season, together with any adult helpers that had been involved in raising brood. Periods of quiet feeding alternate with noisy rapid movement, in follow-my-leader fashion, to a new area; when traversing open areas, flock-members gather together, calling excitedly, before crossing over in single file. Flocks occasionally amalgamate temporarily, with groups of up to 300 individuals reported. May join mixed-species foraging flocks.
Sounds and Vocal Behavior
Very vocal. Usual contact calls a high-pitched , pure, quiet but penetrating “see-see-see” (given especially in flight, particularly when flock in rapid movement, and by individuals isolated from main flock); fuller and flatter, clipped “tup”, usually interspersed with single high, thin “tseet” notes; short, explosive, slurred, rattling trill, falling in pitch towards end, “tsirr” or “tsirrrup”, given in alarm or excitement (frequently without apparent provocation); louder, harder, high-intensity versions given during conflicts between flocks. No territorial song, but occasionally utters quiet twittering and trilling, combining “tup” and “tsirrrup” notes as well as scolds, sometimes in rather structured form (resembling song of an Acrocephalus warbler) and sometimes as a more jumbled collection of notes; this “song” given by both sexes, especially during aggressive encounters with conspecifics, during copulation and nest-building and when feeding young.
Breeding
Season mid-Mar to Jun in Europe and Japan (very exceptionally, fledged young recorded early Feb in England), from Mar onwards in Iran, and Mar–Apr in NE China; single-brooded. Co-operative breeding common; up to eight helpers recorded at single nest, these often adults from failed breeding attempts (male and female of failed pairs do not help at same nest, and, in cases where relationship established, helper is a sibling of male of breeding pair). In late winter or early spring flock breaks up, and unmated females move off to pair with males in adjacent flock. Display includes butterfly-like jerky flight, with presumed male flying up to height of 5–6 m above ground, alternately fanning and closing tail, then diving vertically down; 3–4 birds may display in quick succession or even simultaneously. Each pair sets up loosely defended territory within area occupied by male’s winter flock, but continues to roost with other flock-members until nest so far advanced that it can be used for roosting (usually when dome complete). Nest built by both sexes, over period of up to 33 days, a compact, domed, oval ball of moss, cobweb and hair, covered with up to c. 3000 flakes of lichen (which provide excellent camouflage), average size 160 mm high × 100 mm across, entrance hole at one side near top, lining composed of small feathers (often c. 1500, with maximum of 2680 recorded), adults may continue to add feathers during laying period; placed usually less than 3 m above ground in low thorny bush, but sizable minority sited 6–25 m up in tree fork or against trunk, or hanging among terminal twigs of conifer at up to 35 m above ground, and a few records of nests in hole in ground. Clutch 6–15 eggs, mostly 8–12, average clutch size largest at higher latitudes, eggs white, unmarked or with tiny reddish freckles, 14 × 10·9 mm (nominate race); incubation by female, fed on nest by male, period 12–18 days; chicks cared for by both sexes , assisted by variable number of helpers, presence of which significantly increases breeding success of pair; the provisioning effort of helpers has been found to be positively correlated with their kinship to broods (2); young leave nest after 14–18 days, fed by parents and helpers for at least a further 14 days, family-members then remaining together throughout winter.
Conservation Status
Not globally threatened (Least Concern). Common to fairly common in W Europe and in Korea and Japan, but scarcer towards edge of range in N Scotland and W Ireland and more local and sometimes scarce in E Europe and Siberia, and rare in Kamchatka; in Iran, common in Caspian lowlands but rather scarce in Zagros Mts. Estimated European population more than 3,000,000 breeding pairs, of which c. 30% in Spain; highest recorded densities in Iberia, more than 5000 pairs/50-km square. No major changes in status apparent, although species has expanded N in Norway since c. 1970 and some expansion noted also in N Scotland; in contrast, marked decline in Sweden since 19th century, and similar decline in recent years in Finland, most probably a result of modern forestry practices and the replacement of old-growth habitats with commercial monocultures. Flocks require large winter territories (c. 20–24 ha), and habitat fragmentation and degradation can lead to local extinctions. Notably, very vulnerable to harsh winter weather, following which numbers may decline by up to 80%; can take several years to recover from these setbacks, perhaps owing to high loss of nests (sometimes over 50%) to predators. Success often low, e.g. fledging rate in various studies 16%-28%; main nest predators crows (Corvidae), weasels (Mustela) and snakes.
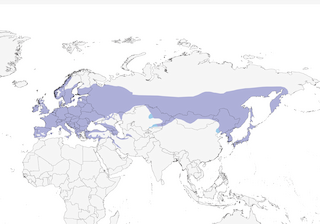
- Year-round
- Migration
- Breeding
- Non-Breeding

















