









European Robin Erithacus rubecula Scientific name definitions
Text last updated January 12, 2013
Sign in to see your badges
Species names in all available languages
| Language | Common name |
|---|---|
| Albanian | Gushëkuqi |
| Arabic | أبو الحناء الأوروبي |
| Armenian | Արշալուսիկ |
| Asturian | Raitñn europñu |
| Azerbaijani | Şəfəq bülbülü |
| Basque | Txantxangorria |
| Bulgarian | Червеногръдка |
| Catalan | pit-roig |
| Chinese (SIM) | 欧亚鸲 |
| Croatian | crvendać |
| Czech | červenka obecná |
| Danish | Rødhals |
| Dutch | Roodborst |
| English | European Robin |
| English (United States) | European Robin |
| Faroese | Bringureyði |
| Finnish | punarinta |
| French | Rougegorge familier |
| French (France) | Rougegorge familier |
| Galician | Paporrubio común |
| German | Rotkehlchen |
| Greek | Κοκκινολαίμης |
| Hebrew | אדום חזה |
| Hungarian | Vörösbegy |
| Icelandic | Glóbrystingur |
| Italian | Pettirosso |
| Japanese | ヨーロッパコマドリ |
| Korean | 꼬까울새 |
| Latvian | Sarkanrīklīte |
| Lithuanian | Liepsnelė |
| Mongolian | Өрнийн зээрд омруут |
| Norwegian | rødstrupe |
| Persian | سینه سرخ اروپایی |
| Polish | rudzik |
| Portuguese (Portugal) | Pisco-de-peito-ruivo |
| Portuguese (RAA) | Vinagreira |
| Portuguese (RAM) | Papinho |
| Romanian | Măcăleandru |
| Russian | Зарянка |
| Serbian | Crvendać |
| Slovak | červienka obyčajná |
| Slovenian | Taščica |
| Spanish | Petirrojo Europeo |
| Spanish (Spain) | Petirrojo europeo |
| Swedish | rödhake |
| Turkish | Kızılgerdan |
| Ukrainian | Вільшанка |
Erithacus rubecula (Linnaeus, 1758)
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
The Key to Scientific Names
Legend Overview
Originally Appeared in

Field Identification
14 cm; 14–25 g.Nominate subspecies is olive-brown above , with orange face and breast fringed by band of pale blue-gray on neck side to breast side; buff lower flanks and white belly to vent; bill blackish, legs pinkish-brown. Sexes similar. Juvenile is extensively mottled brown and buff. Subspecies melophilus is warmer and darker above, deeper orange-rufous on face and breast, darker buff flanks; witherbyi is like previous but smaller; superbus is dark grayish-olive above, wider ash-gray band from crown side to breast side, still deeper orange-rufous below, belly and vent whiter, with somewhat different wing shape; valens is slightly paler than nominate, with rufous uppertail-coverts and tail base; caucasicus is like nominate but slightly browner above, with rufous uppertail-coverts and tail base; hyrcanus is longer-billed, browner above, rufous-orange below, with rufous-chestnut uppertail-coverts; tataricus is paler and grayer above, paler orange below.
Systematics History
Editor's Note: This article requires further editing work to merge existing content into the appropriate Subspecies sections. Please bear with us while this update takes place.
Has been suggested that Canary Island subspecies superbus merits treatment as a separate species, with birds on Gran Canaria proposed as a separate subspecies (“marionae”) on grounds of considerable genetic isolation (1, 2); this species status, however, is based on very minor levels of differentiation, and taxonomic distinction between the two populations probably not justified without confirmatory differences in morphological characters. Geographical variation extremely slight, with considerable intergradation among western subspecies; valens and caucasicus sometimes merged with hyrcanus. Other described subspecies are armoricanus (described from northwestern France), sardus (Sardinia) and balcanicus (western Turkey), all synonymized with nominate. Nine subspecies recognized.
Subspecies
Erithacus rubecula melophilus Scientific name definitions
Distribution
British Isles; non-breeding also southwestern Europe.
Erithacus rubecula melophilus Hartert, 1901
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- melophilus
- Melophilus
The Key to Scientific Names
Legend Overview
Erithacus rubecula rubecula Scientific name definitions
Distribution
Western Europe, northwestern Morocco, Azores, Madeira and western Canary Islands.
Erithacus rubecula rubecula (Linnaeus, 1758)
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
The Key to Scientific Names
Legend Overview
Erithacus rubecula tataricus Scientific name definitions

Distribution
Urals and southwestern Siberia; non-breeding southwestern Asia.
Erithacus rubecula tataricus Grote, 1928
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- tatarica / tataricus
The Key to Scientific Names
Legend Overview
Erithacus rubecula superbus Scientific name definitions

Distribution
Central Canary Islands (Tenerife).
Erithacus rubecula superbus Koenig, 1889
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- superba / superbus
The Key to Scientific Names
Legend Overview
Erithacus rubecula marionae Scientific name definitions
Distribution
Central Canary Islands (Gran Canaria).
Erithacus rubecula marionae Dietzen et al., 2015
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- marionae
The Key to Scientific Names
Legend Overview
Erithacus rubecula witherbyi Scientific name definitions
Distribution
Northern Algeria and northern Tunisia.
Erithacus rubecula witherbyi Hartert, 1910
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- witherbyi
The Key to Scientific Names
Legend Overview
Erithacus rubecula valens Scientific name definitions
Distribution
Southern Crimea.
Erithacus rubecula valens Portenko, 1954
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- valens
The Key to Scientific Names
Legend Overview
Erithacus rubecula caucasicus Scientific name definitions
Distribution
Eastern Turkey east to the Caucasus region.
Erithacus rubecula caucasicus Buturlin, 1907
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- caucasica / caucasicus
The Key to Scientific Names
Legend Overview
Erithacus rubecula hyrcanus Scientific name definitions
Distribution
Southeastern Azerbaijan and northern Iran; non-breeding also Middle East.
Erithacus rubecula hyrcanus Blanford, 1874
Definitions
- ERITHACUS
- erithacus
- rubecula
- Rubecula
- hyrcana / hyrcanus
The Key to Scientific Names
Legend Overview
Distribution
Editor's Note: Additional distribution information for this taxon can be found in the 'Subspecies' article above. In the future we will develop a range-wide distribution article.
Habitat
Forest undergrowth and edge, preferring conifer tracts in some parts of range and deciduous woodland in others; copses and adjacent open land, farmland woodlots, thickets along watercourses, hedgerows with some tall trees, orchards,gardens and parks; key requirements for long-term site-occupancy are cool shade, medium-height cover with perches, and patches of bare ground. In urban areas in parts of Europe. Breeding habitat in northwestern Africa mainly montane forest (Atlas cedar and oak) with thick undergrowth (Viburnum, Ilex, Erica), humid leaf litter and dead wood, sparsely at 700–1000 m, commonly 1000–2000 m, rarely higher. Non-breeding migrants in North Africa and Middle East occupy woodland, farmland, dense macchia, large gardens, orchards and plantations. Keeps to open undergrowth.
Movement
Poorly understood. In general, populations east of line from Norway and Sweden south to Central Europe are full migrants, moving south to Mediterranean Basin, Black Sea hinterland, S Caspian, Mesopotamia and Gulf region (although subspecies caucasicus and hyrcanus are partial and/or vertical migrants); those west of line from Germany to Balkans partially migratory or largely resident (sedentary on Canary Islands, with little vertical movement). In British Isles partial migration occurs (main passage mid-Aug to end Oct, peak early Oct), as indicated by male bias in wintering population, suggesting that females migrate (69–77% of males in four study areas sedentary), but distribution of migrants uncertain, although most seem to move short distances and fewer than 5% emigrate; spring passage less notable and extending late Feb to May. Birds from Northern Europe wintering in Spain keep mainly to south and east (especially Balearic Islands), with 75% of birds immigrating mid-Oct to mid-Nov. Individuals ringed in Poland migrate to broad area of Mediterranean, from Iberian Peninsula east to Balkans; autumn passage on northern Polish coast mid-Aug to early Nov, peak late Sept, although birds in northern Norway linger into early Oct, and those in Russia (Urals, Moscow) usually vacate breeding areas late Sept to mid-Oct. Scandinavian and Russian birds of nominate subspecies appear to winter in western half of Europe and North Africa. Birds ringed in Belgium recovered 2400 km south (northern Saharan fringes) and 1700 km north, average speed during long-distance displacements 100 km/day; most Belgian birds winter in southwestern Europe, and about half of Belgian wintering population composed of immigrants. One study in Switzerland found that first-years start their passage c. 20 days before adults. In Italy autumn stopovers longer than spring ones. Resident northwestern Africa, and no evidence of altitudinal movement. Also abundant winter visitor to North Africa; autumn passage of Northern European (and possibly Mediterranean) birds from late Sept (sometimes earlier), peak Oct, birds present in region until Mar; records at Saharan oases mostly late Dec to Mar. Passage in Syria from end Oct and in Mar; in Israel mainly Nov and Mar. Spring passage in North Africa starts Feb, with peak mid-Mar to early Apr, last leaving by late Apr (earlier in west than in east); Cyprus largely vacated by mid-Apr, and some higher sites in Alps not reoccupied until early May, about same time as return to northern Scandinavian sites.
Diet and Foraging
Invertebrates, fruits and seeds; also very small vertebrates (including fish and lizards), carrion , left-overs and birdtable foods.Invertebrates include adult and larval beetles of many families, adult and larval flies of many families, ants, bees, sawflies, crickets, grasshoppers, bugs, earwigs, thrips, adult and larval butterflies and moths; also spiders, mites, woodlice, sandhoppers, millipedes, centipedes, small molluscs, earthworms. Fruits and seeds include juniper (Juniperus), yew (Taxus), spurrey (Spergula), strawberry (Fragaria), apple (Malus), cherries (Prunus), cotoneaster (Cotoneaster), oleander (Olea), pistachio (Pistacia), spindle (Euonymus), sea buckthorn (Hippophae) buckthorn (Rhamnus), rowan (Sorbus), bramble (Rubus), current (Ribes), bilberry (Vaccinium), elder (Sambucus), viburnum (Viburnum), honeysuckle (Lonicera), dogwood (Cornus), oak (Quercus), vine (Vitis), wild vine (Ampelopsis), privet (Ligustrum), mistletoe (Viscum) and nightshade (Solanum). In southern Spain, sample of birds in holm oak (Quercus ilex) woods in winter (Oct–Feb) yielded 1900 invertebrates, of which 76% ants, 12% beetles, and 12% larvae and earwigs Forficula, but birds also took pieces of acorn (varied monthly from 16·5% by volume to as much as 78%); sample of birds in farmland (Nov–Jan) yielded 300 invertebrates, of which 73% ants, 15% beetles and the rest berries (26–42% by volume); and at highland locality fruits of Pistacia lentiscus formed 31% of plant diet, Phillyrea latifolia 19% and Viburnum tinus 6%, while at lowland site Pistacia lentiscus rose to 89%. In another study in Mediterranean scrub, southern Spain, fleshy fruits of nine plants (P. lentiscus most important) plus acorn endosperm varied 53–77·5% in diet volume over three winters, and at least 89% of individual diet samples contained at least some fruit; monthly averages of fruit volume varied inversely with arthropod abundance, but reliance on food apparently a response to fruit availability (not arthropod scarcity), and body condition of birds unaffected by different levels of fruit consumption. In Germany, individuals in hedgerow habitats took 60% beetles, supplemented by flies, millipedes and spiders; majority of prey small, 2–6 mm in length. Main foods of spring migrants in Russian Baltic were beetles, ants, myriapods and spiders; in Crimea in summer, stomachs contained 39% beetles, 16% hymenopterans (majority ants), 15% millipedes, 8% bugs, 6% caterpillars, among others; in Armenia in winter, stomachs held beetles (of at least seven families), bugs, ants, caterpillars and plant material. Two main foraging methods: perching on low lookout post (bush, branch, fence) in open or partially open areas and flying down to take prey from ground, returning to same or similar perch; and pursuing and taking prey in hopping gait on ground in sheltered vegetation (this strategy increasing in colder weather). Occasionally sallies after flying insects. In Northern Europe a frequent visitor to birdtables, where very fond of bread . Will follow animals such as Ring-necked Pheasant (Phasianus colchicus) and mole (Talpa) for invertebrates disturbed by their passage; renowned for habit of accompanying gardeners digging soil. Takes items from streams and margins of pools and creeks, sometimes foraging in intertidal zones; recorded as catching insects by artificial light. Male strongly territorial all year; female defends separate area in winter. Size of winter territory in Britain 0.43 ha, in Israel 0.6–1 ha; in North Africa c. 300 birds in 1 km² indicates similar values.
Sounds and Vocal Behavior
Song , all year, a series of highly variable phrases, each consisting of thin high silvery whistling that typically mixes a short melodious warbled passage and several long-drawn but fading notes, rarely with some mimicry. Song in autumn to late winter (by both sexes) differs from that in spring (almost entirely by male) in being quieter, with longer phrasing and more melancholy tone; songs during fights have more strangled, slurred quality, and often repeat same phrases. Birds in Canary Islands have somewhat simpler songs. Subsong (both sexes) a subdued complex rambling warble, incorporating more obvious mimicry than full song. Calls include irregularly repeated staccato “tic” in excitement, often associated with territorial defense (can become rapid crackling tittering in greater excitement: “tic, tic, tikeritititit, tikerititititititit, tic, tic…”); thin “tswiiii” in contact-alarm; high sharp thin “tsiiip” or “sssiiip” in alarm; high piercing “siii” in aggression; soft high “dziip” for contact and by food-begging female; hissing high “seep” or “sweezeezeez” by female as being fed, with similar call during copulation; thin metallic “peep” when foraging; soft sibilant “sip” or “sissip” from nocturnal migrants; and various churrs.
Breeding
Mainly early Apr to mid-Jun in British Isles; end Apr to late Jul in Central Europe; from mid-May in northern Russia and mid-Apr in south; late Apr to early Aug in southern Caucasus region (Armenia); mid-Mar to Jun in Canary Islands; May–Jul in Morocco and Apr–May in Algeria; single-brooded in north of range, elsewhere two broods, rarely three. Breeding territory in Britain average 0.84 ha, in Canary Islands 0.9–1.5 ha. Nest a cup of moss, grass, leaves and twigs, lined with fine grass and hair, placed on bulky mat of leaves and always sited in recess, usually in low undergrowth, hedge, grass bank, wall, rock face, tree cavity , tree roots, nestbox , usually low down, not above 5 m and fairly often on ground under tussock; very fond of ivy-clad walls and tree trunks for nesting. Eggs 4–7 (4–5 in Algeria), whitish or bluish-white (whitish-pink in subspecies superbus) with small reddish spots; incubation period 12–21 days, mostly c. 14 days; nestling period 10–18 days, generally c. 13 days; post-fledging dependence 16–24 days. In Britain, 71% of 1,400 eggs and 77% of hatchlings fledged, giving overall success 55% (overall success 53% in Mar–Apr, 61% in May, 46% in Jun–Jul). In Britain, annual adult mortality 62%, yearling mortality from 1st Aug 72%; annual overall mortality in Finland 76%, in Europe as a whole 58–62%; mortality rates from Belgian ringing data 0.71 in first year and 0.54 after first year (only 1% of birds recovered after fourth calendar year); causes of mortality of ringed birds in northwestern Europe are domestic predator 50%, human-related (accidental) 32%, human-related (deliberate) 4%, other 14%. Age of first breeding 1 year. Recorded longevity more than 8 years.
Conservation Status
Not globally threatened. European population in mid-1990s judged stable and estimated at 32,387,823–42,974,703 pairs, with estimated additional 10,000,000–100,000,000 pairs in Russia (apparently west of Urals) and 10,000–100,000 pairs in Turkey; population in Spain then assessed at 1,200,000–3,000,000 pairs and probably stable or growing. By 2000, total European population (including European Russia and Turkey) revised to 43,000,000–83,000,000 pairs and considered stable. Densities in Europe may reach 1 pair/ha (100 pairs/km²) in good woodland habitat and suburban gardens, but drop to one-third of this level in farmland with small fields and hedgerows. Relatively scarce resident in NW Africa; 25·8 pairs/km² in Morocco. Often abundant in winter in North African coastal zone, e.g. 62 birds/km² in hillside macchia and 250/km² in mixed woodland in Algeria, and 130/km² in mixed countryside in Morocco; also frequent to common in Libya and along Egyptian coast to western Sinai, where up to 50/km²; scarce south of coastal zones. Winter density of 2.6 birds/ha (260/km²) found in semi-natural woodland in city park in Rome (Italy), very close to the figure for mixed woodland in Algeria. No pressing conservation issues, but many individuals still caught and killed for food in Mediterranean Basin in winter.

- Year-round
- Migration
- Breeding
- Non-Breeding

eBird and Macaulay Library
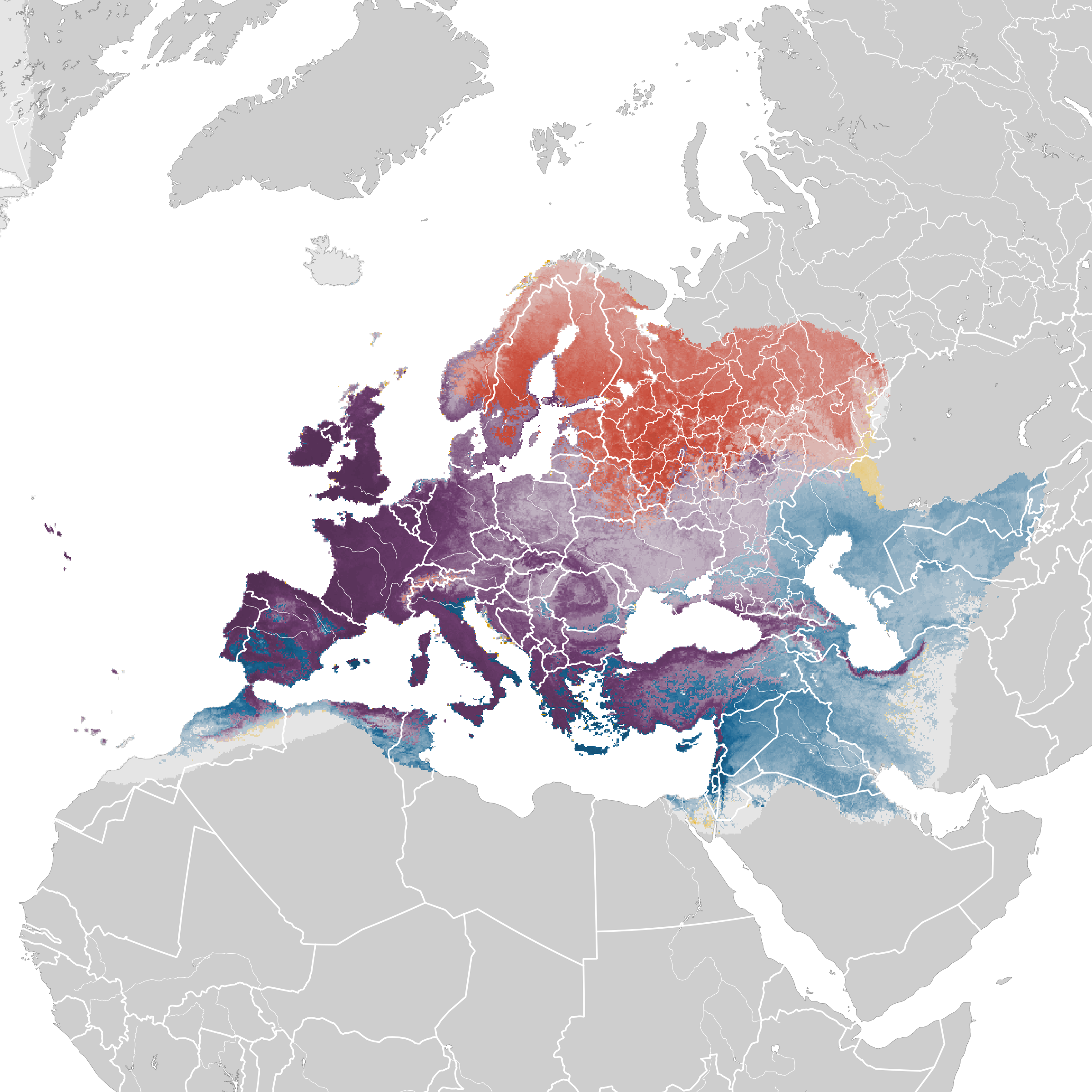
eBird Status and Trends

















